Кольчатые черви обладают самой высокой организацией по сравнению с другими типами червей; у них впервые появляются вторичная полость тела, кровеносная система, более высоко организована нервная система. У кольчатых червей внутри первичной полости образовалась другая, вторичная полость с собственными эластичными стенками из клеток мезодермы. Её можно сравнить с подушками безопасности, по паре в каждом членике тела. Они «раздулись», заполнили пространство между органами и поддерживают их. Теперь каждый сегмент получил свою собственную опору из мешков вторичной полости, наполненных жидкостью, а первичная полость эту функцию утратила.
Обитают в почве, пресной и морской воде.
Внешнее строение
Дождевой червь имеет почти круглое в поперечном разрезе тело длиной до 30 см; насчитывают 100-180 сегментов, или члеников. В передней трети тела находится утолщение — поясок (его клетки функционируют в период полового размножения и откладки яиц). По бокам каждого сегмента развиты по две пары коротких упругих щетинок, которые помогают животному при передвижении в почве. Тело имеет красновато-коричневую окраску, светлее на плоской брюшной и темнее на выпуклой спинной стороне.
Внутреннее строение
Характерной особенностью внутреннего строения является то, что у дождевых червей развиты настоящие ткани. Снаружи тело покрыто слоем эктодермы, клетки которой образуют покровную ткань. Кожный эпителий богат слизистыми железистыми клетками.
Мышцы
Под клетками кожного эпителия находится хорошо развитая мускулатура, состоящая из слоя кольцевых и находящегося под ним более мощного слоя продольных мышц. Мощные продольные и кольцевые мышцы меняют форму каждого членика отдельно.
Дождевой червь попеременно то сжимает и удлиняет их, то расширяет и укорачивает. Волнообразные сокращения тела позволяют не только ползти по норке, но и раздвигать почву, расширяя ход.
Пищеварительная система
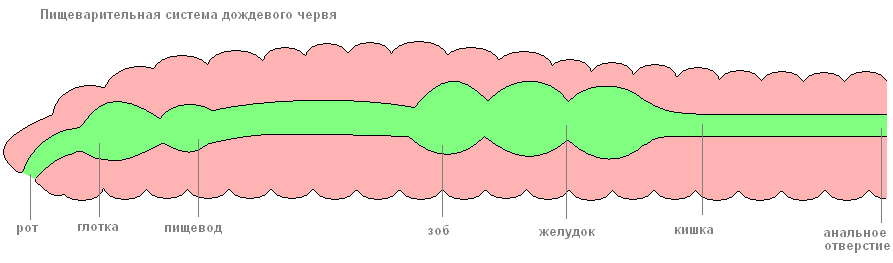
Пищеварительная система начинается на переднем конце тела ротовым отверстием, из него пища поступает последовательно в глотку, пищевод (у дождевых червей в него впадают три пары известковых желёз, поступающая из них в пищевод известь служит для нейтрализации кислот гниющих листьев, которыми питаются животные). Затем пища переходит в расширенный зоб и небольшой мускульный желудок (мышцы в его стенках способствуют перетиранию пищи).
От желудка почти до заднего конца тела тянется средняя кишка, в которой под действием ферментов пища переваривается и всасывается. Непереваренные остатки поступают в короткую заднюю кишку и выбрасываются наружу через заднепроходное отверстие. Дождевые черви питаются полусгнившими остатками растений, которые они глотают вместе с землёй. При прохождении по кишечнику почва хорошо перемешивается с органическими веществами. Экскременты дождевых червей содержат в пять раз больше азота, в семь раз больше фосфора и в одиннадцать раз больше калия, чем обычная почва.
Кровеносная система

Кровеносная система замкнутая, состоит из кровеносных сосудов. Вдоль всего тела над кишечником тянется спинной сосуд, а под ним — брюшной.
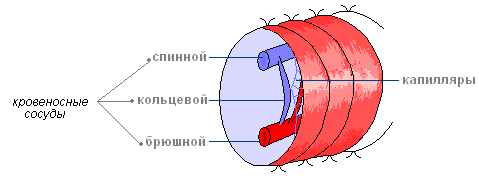
В каждом сегменте их объединяет кольцевой сосуд. В передних сегментах некоторые кольцевые сосуды утолщены, их стенки сокращаются и ритмически пульсируют, благодаря этому кровь перегоняется из спинного сосуда в брюшной.
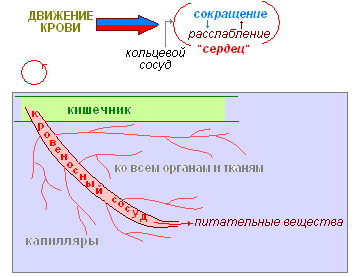
Красный цвет крови обусловлен наличием в плазме гемоглобина. Он играет ту же роль что и у человека — питательные вещества растворёнными в крови разносятся по всему телу.
Дыхание
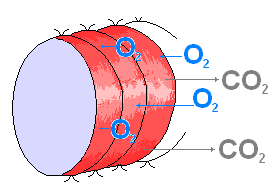
Для большинства кольчатых червей, в том числе и дождевых, характерно кожное дыхание, практически весь газообмен обеспечивается поверхностью тела, поэтому черви очень чувствительны к влажной почве и не встречаются в сухих песчаных почвах, где их кожа скоро подсыхает, а после дождей, когда в почве много воды, выползают на поверхность.
Нервная система

В переднем членике червя находится окологлоточное кольцо — самое крупное скопление нервных клеток. С него начинается брюшная нервная цепочка с узлами нервных клеток в каждом членике.

Такая нервная система узловатого типа образовалась при слиянии нервных тяжей правой и левой стороны тела. Она обеспечивает самостоятельность члеников и слаженную работу всех органов.
Органы выделения
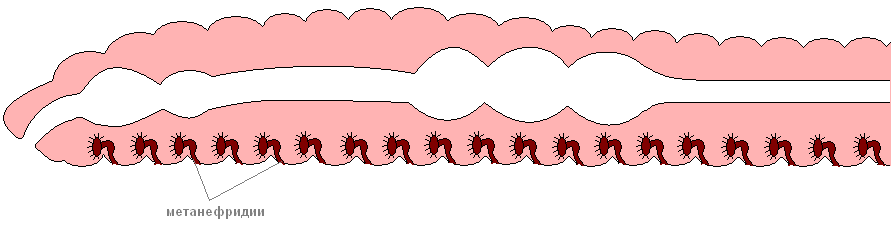
Органы выделения имеют вид тоненьких петлеобразно изогнутых трубочек, которые одним концом открываются в полость тела, а другим наружу. Новые, более простые воронкообразные органы выделения — метанефридии выводят вредные вещества во внешнюю среду по мере их накопления.

Размножение и развитие
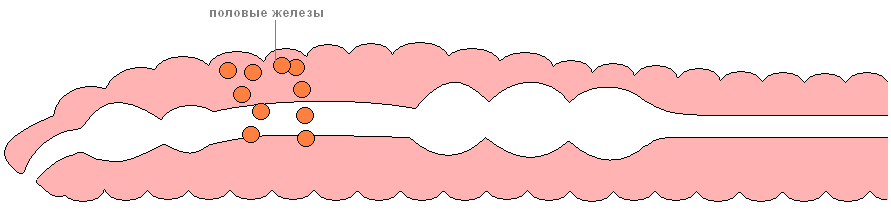
Размножение происходит только половым путём. Дождевые черви — гермафродиты. Половая система их расположена в нескольких сегментах передней части. Семенники лежат впереди яичников. При спаривании сперматозоиды каждого из двух червей переносятся в семяприёмники (особые полости) другого. Оплодотворение червей перекрёстное.

Во время копуляции (спаривание) и откладки яиц клетки пояска на 32-37 — сегменте выделяют слизь, служащую для образования яйцевого кокона, и белковую жидкость для питания развивающегося зародыша. Выделения пояска образуют своеобразную слизистую муфту (1).
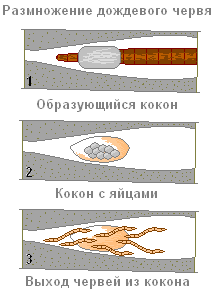
Червь выползает из неё задним концом вперёд, откладывая в слизь яйца. Края муфты слипаются и образуется кокон, который остаётся в земляной норке (2). Эмбриональное развитие яиц происходит в коконе, из него выходят молодые черви (3).
Органы чувств
Органы чувств развиты очень слабо. У дождевого червя нет настоящих органов зрения, их роль выполняют отдельные светочувствительные клетки, находящиеся в кожном покрове. Там же помещаются рецепторы осязания, вкуса, обоняния. Дождевые черви способны к регенерации (легко восстанавливает заднюю часть).
Зародышевые листки
Зародышевые листки основа всех органов. У кольчатых червей эктодерма (наружный слой клеток), энтодерма (внутренний слой клеток) и мезодерма (промежуточный слой клеток) появляются в начале развития как три зародышевых листка. Они дают начало всем основным системам органов, включая вторичную полость и кровеносную систему.
Эти же системы органов сохраняются в дальнейшем у всех высших животных, причём они формируются из тех же трёх зародышевых листков. Так высшие животные в своём развитии повторяют эволюционное развитие предков.
Многие люди недооценивают значимость работы земляных дождевых червей. Эти представители беспозвоночного царства известны прежде всего тем, что в больших количествах выползают из земли после сильного дождя. Нередко используются они и как приманка многочисленными любителями рыбалки. Еще Дарвин отметил тот факт, что черви выполняют важную функцию в природе, выступая в роли своеобразных агротехников. В процессе создания разветвленной системы тоннелей, которую прорывает дождевой червь, образуется отличная аэрация посредством поступления воздуха к внутренним слоям грунта.
Благодаря превосходной аэрации облегчается дыхательная деятельность многих растений. Питаясь органикой и отходами, черви обеспечивают измельчение почвенных компонентов, обогащая их при этом своими выделениями. Удивительная способность представителей данного вида состоит в возможности обеззараживать огромные участки почвы, стерилизуя ее от вредоносных бактерий. Благодаря бесчисленным норам, образующим подобие капиллярной системы, обеспечивается идеальный дренаж и проветривание грунта.
Тело дождевого червя в длину может достигать трех метров. Однако на территории России встречаются преимущественно особи, дина тела которых не превышает 30 сантиметров. Для того, чтобы передвигаться, червь применяет небольшие щетинки, которые находятся на разных частях туловища. В зависимости от разновидности, сегментов может быть от 100 до 300. Кровеносная система является замкнутой и развита весьма неплохо. Составляют ее одна артерия и одна центральная вена.
Строение дождевого червя – весьма необычное. Дыхание реализуется при помощи особых сверхчувствительных клеток. Кожный покров продуцирует защитную слизь с достаточным количеством натуральных антисептиков. Строение мозга довольно примитивное и включает в себя всего лишь два нервных узла. Земляные черви по итогам лабораторных экспериментов подтвердили свои выдающиеся способности к регенерации. Отрубленный хвост отрастает заново спустя небольшой отрезок времени.
Половые органы дождевого червя тоже устроены весьма необычно. Каждая особь представляет собой гермафродита. Обладает она и мужскими органами. По биологическим факторам всех подобных червей можно разделить на несколько подгрупп. Представители одной из них занимаются поисками пропитания на поверхности почвенного слоя. Другие используют в пищу непосредственно сам грунт и показываются из-под земли крайне редко.
Земляной червь относится к типу кольчатых. Под кожным слоем находится развитая система мускулов, состоящая из мышц различной формы. Ротовое отверстие, из которого пища попадет в пищевод через глотку, находится на передней части туловища. Оттуда она транспортируется в область расширенного зоба и небольших размеров мускульного желудка.

Роющие и подстилочные дождевые черви обитают в местах с рыхлой и увлажненной почвой. Предпочтение отдается влажным грунтам субтропиков, болотистым землям и берегам различных водохранилищ. На степных территориях обыкновенно встречаются почвенные разновидности червей. В тайге и лесотундре проживают подстилочные виды. Наибольшей концентрацией особей может похвастать хвойная широколиственная полоса.
Какую почву любят черви?
Почему дождевые черви обожают почвы супесчаного типа и суглинки? Подобный грунт отличается низкой кислотностью, которая самым лучшим образом подходит для их жизнедеятельности. Уровень кислотности выше pH 5,5 является губительным для организмов этих представителей кольчатого типа. Влажные почвы – одно из обязательных условий для расширения популяции. На время засушливой и жаркой погоды черви уходят глубоко под землю и теряют возможность размножаться.
Характер и образ жизни дождевого червя
Активная и продуктивная жизнь дождевого червя выпадает на темное время суток. Как только наступает ночь, множество особей выползает на поверхность грунта в поисках пропитания. Однако хвост, как правило, остается в земле. К утру они возвращаются в свои норы с добычей, затаскивая в них кусочки пищи и маскируют вход в свое убежище посредством травинок и листвы.

Роль дождевых червей в природе сложно оценить чрезмерно. Червь в буквальном смысле пропускает сквозь себя невероятное количество грунтовой смеси, обогащая ее полезными ферментами и убивая вредные вещества и бактерии. Передвигается червь посредством ползания. Втягивая один конец тела и цепляясь щетинками за шероховатости земли, он подтягивает заднюю часть, прокладывая подобным образом свои многочисленные ходы.
Как дождевые черви переживают зиму?
На зимний период абсолютное большинство особей впадает в спячку. Резкое понижение температуры способно моментально уничтожить червей, поэтому они стараются заблаговременно зарыться в почву на глубину, нередко превышающую один метр. Дождевые черви в почве выполняют наиважнейшую функцию по ее естественному обновлению и обогащению различными веществами и микроэлементами.
Польза
В процессе переваривания полуферментированных листьев организм червей вырабатывает специфические ферменты, которые способствуют активной генерации гумусовой кислоты. Почва, подвергшаяся разрыхлению земляными червями, является оптимальной для самых разнообразных представителей растительного царства. Благодаря системе запутанных тоннелей обеспечивается превосходная аэрация и вентиляция корней. Таким образом передвижение дождевого червя является важным фактором в задаче восстановления полезных качеств почвы.
Дождевой червь по факту очень полезен для человека. Он делает слои почвы плодородными и обогащает их всевозможными питательными веществами. Однако общее количество особей во многих регионах России стремительно сокращается. Происходит это по причине бесконтрольного внесения ядохимикатов, удобрений и минеральных смесей в грунт. Также на земляных червей охотятся многочисленные птицы, кроты, и разные грызуны.
Чем питаются дождевые черви?
Ночью дождевой червь выползает на поверхность и стягивает полусгнившие остатки растений и листьев в свое убежище. Также в его рацион питания входит почва, богатая перегноем. Один представитель вида может переработать до половины грамма почвы за сутки. Учитывая, что на площади в один гектар может находиться одновременно до нескольких миллионов особей, они способны выступить в качестве незаменимых преобразователей грунта.
После дождя на асфальте и поверхности почвы можно увидеть большое количество червей, что же заставляет их вылезать наружу? Даже название «дождевые черви» указывает на то, что они очень любят влагу и активизируются после дождя. Рассмотрим несколько возможных причин почему дождевые черви вылезают после дождя на поверхность земли.

Температура почвы
Есть мнение, что черви вылезают на поверхность в поисках тепла, так как после дождя температура почвы понижается на несколько градусов, что вызывает дискомфорт для них.
Изменение кислотно-щелочного баланса
Другая теория говорит, что червяки вылезают на поверхность из-за изменения кислотно-щелочного баланса почвы после дождя, она становится более кислой, что негативно влияет на этих землекопов. По мнению исследователей, экстренная эвакуация на поверхность почвы спасает их от гибели в кислой среде.
Недостаток воздуха
Третья теория объясняет, что после дождя в верхнем слое почве становится больше кислорода, поэтому червяки массово вылезают наверх. Вода обогащает верхние слои земли кислородом, а многие виды червей любят влагу и жизненно нуждаются в достаточном количестве кислорода. А через поверхность тела кислород поглощается лучше всего именно во влажной среде.
Путешествия
Британский ученый Крис Лоу предположил, что черви вылезают на поверхность земли во время дождя для того, чтобы совершить протяженное путешествие на новую территорию. По поверхности черви способны проползти намного дальше, чем под землей, а сухая почва вызывает дискомфорт при перемещении, создается сильное трение, к поверхности тела червяка прилипают песчинки, ранящие его. А после дождя поверхность земли сильно увлажнена, что позволяет им беспрепятственно путешествовать на новые участки грунта.
Шум дождя
Другой ученый, профессор Джозеф Горрис из США, предположил, что земляные черви пугаются шума дождя, так как вибрация, которые он создает, похожи на звук приближения их главного врага — крота. Именно поэтому некоторые рыбаки используют прием для выманивания приманки на поверхность: вставляют в землю палку, на ее поверхности закрепляют лист железа и дергают его так, чтобы создать вибрацию, коротая передастся в землю через палку. Пугаясь, черви выбираются на поверхность земли и становятся легкой добычей для опытных рыбаков.
Размножение и продолжительность жизни дождевых червей
Дождевой червь является гермафродитом. Имеет он как женские, так и мужские половые органы. Однако на самооплодотворение он не способен. С наступлением требуемых для размножения теплых климатических условий особи сползаются попарно, прикладываясь друг к другу брюшной областью, и производят своеобразный обмен семенем. После происходит преобразование муфты в кокон, в котором и развиваются яйца.

Некоторые разновидности отличаются бесполым размножением. Тело червя разделяется надвое, при этом одна из частей регенерирует передний конец, а другая – задний. Также существуют виды червей, размножающихся без семяприемников посредством откладывания сперматофор. Продолжительность жизни червей может превышать десять лет.
Ещё с давних времён, человечество обратило особое внимание на таких неприглядных созданий, как дождевой червь. А учёные умы, в лице Чарльза Дарвина, спустя десятилетия, долгие годы изучали их строение и значение в земледелии. И недаром. Ведь с наступлением весеннего тепла, дождевые черви приступают к кропотливой работе и трудятся, сами того не ведая, на благо людям.
Особенности и среда обитания
Дождевой червь, он же кольчатый – всем известный житель на любом приусадебном участке. И казалось бы, абсолютно незаметное, никому не нужное создание.
Однако, любой человек, хоть как то связанный с землей, будет очень рад таким жителям его огорода. В Российской Федерации существует не более сотни видов дождевого червя. А вот по всему миру их полторы тысячи разновидностей.
Принадлежит он к семейству кольчатых червей, малощетинкового класса. Всё его длинное тельце состоит из множества колец. Их может быть семьдесят, а может и все триста. Так как в длину он вырастает более двадцати пяти сантиметров.
Но есть и самые маленькие, двух-трёх сантиметровые. В Австралийские земляные черви, достигают двух с половиной метров в своих размерах. Его цвет в прямом смысле слова серо-буро — малиновый.
Так же, на каждом кольце, или его ещё называют сегмент, находятся щетинки. У обычных наших огородных червяков, как правило, щетинок вырастает по восемь штук. Они классифицируются как малощетинковые.
Однако есть и тропические, многощетинковые виды червей, у которых ворсинки растут десятками. Щетинки помогают червям ползать, абсолютно по всем почвенным бугоркам или зарываться в норы.
Обнаружить их можно, взяв червяка в руки и проведя пальцем сзади наперёд. Но так как, неопытному человеку, тяжело определить, где у него зад, можно просто легонько провести рукой вдоль тела и обратно. Сразу можно ощутить. В одну сторону червяк будет абсолютно гладким, а проведя в противоположную – шероховатым.

Кто хоть раз брал червяка в руки, знает, что он весь покрыт не очень приятной слизью, которая ему жизненно необходима. Во-первых – слизь помогает беспозвоночному свободно передвигаться в земле. Во вторых – так как у червя нет лёгких, дышит он через кожу. И благодаря влаге, находящейся на слизи, тело насыщается кислородом.
Само тело дождевого червя, состоит их двух групп мышечной ткани. Они продольные и поперечные. Поперечные мышцы, находятся под защитным верхним слоем кожи червя.
С их помощью червь становится максимально длинным. А более сильные мышцы – продольные. Они сокращают, уменьшают тело. Вот так, то удлиняясь, то укорачиваясь, животное и передвигается.
Дождевой червь относится к вторичнополостным животным. Следовательно, у него есть полноценная замкнутая система кровообращения. Так как жизнедеятельность у них активная.
Мышцы сокращаются во много раз чаще, чем у червей первичнополостных. Для этого им нужна кровь, чтобы обеспечивать червя всеми питательными веществами и кислородом.
В строении дождевого червя есть пара кровеносных сосудов, один их них называется спинным, второй брюшным. Кольцевые сосуды, соединяют их между собой. Кровь протекает по ним с заду на перёд, и наоборот.
В каждом кольце, или как оно ещё называется – сегменте, есть пара трубочек. Воронки, имеющиеся на их концах, открываются и через низ выводятся испражнения дождевого червя. Это принцип работы выделительной системы.

Что же касается нервной системы, она узловая. Её составляющие – брюшная нервная цепочка и окологлоточное нервное кольцо. Эти окончания состоят из волокон, а те в свою очередь, отвечают на позывы сокращённых мышц червя. Благодаря им, червь может кушать, целенаправленно двигаться, размножаться, развиваться.
В строении органов дождевого червя, отсутствуют те, которые отвечают за обоняние, осязание, зрение, ощущение. Но имеются определённые клетки, они располагаются вдоль всего тела беспозвоночного. С их помощью червь ориентируется в темноте и непроходимой земле.
Характер и образ жизни
Ещё Чарльз Дарвин предположил наличие интелекта у дождевых червей. Наблюдая за ними, он заметил, что затаскивая к себе в жилище сухой листок, тот повернут именно узкой стороной. Что облегчает проходимость листика через плотную, землянистую нору. А вот иглы еловых наоборот, берёт за основание, чтобы они не раздваивались.
Весь день, вся жизнь дождевого червя расписан по минутам. Он то и дело лазит в земле, делает ходы, проглатывая её. Роет норы червяк двумя способами. Он или, как уже говорилось, проглатывает землю, постепенно продвигаясь вперёд.
В случае если земля слишком твёрдая. И следом оставляя свои биологические отходы. Или, распихивает её своим утончённым концом, в разные стороны, и делает себе ходы. Проходы получаются наискось вертикальные.

Тек же, дождевой червь, промышляя в почве , затаскивает к себе в норы, для утепления, различные листики, жилки с листочков, тонкие бумажечки и даже клочки шерсти. Его норы глубиной до одного метра. А у червей покрупнее размерами, и все десять метров. Трудится червь в основном в ночные часы.
А почему же дождевые черви огромными количествами выползают на поверхность. Это значит, что ему нечем дышать. Такое обычно случается после обильных дождей. Земля забивается влагой, и ему совсем нет кислорода. По приходу холодов, дождевой червь уходит глубоко в почву.
Питание дождевого червя
Питание у червя достаточно типичное. Заглатывая в больших количествах землю вместе с пищей. Для еды им годятся привялые и немного подгнившие листочки, грибочки. Но она не должна иметь неприятного запаха, иначе червь её есть не будет.
Оказывается, дождевые черви, даже сооружают себе целые кладовые комнаты, и складывают туда к зиме еду. Кушают они её только в случае критической необходимости. Например, в зимнее время, когда грунт полностью промёрзший, и ни о какой наземной пище не может быть и речи.
Всосав пищу вместе с комом земли, через глотку, мышечными движениями, то расширяя своё тело, то сужая, он заталкивает её к задней части пищевода в зоб. После, она проникает в желудок. Из желудка отправляется перетравливаться в кишечнике, благодаря ферментам, выходит на-наружу полезнейшими биомассами.

Проделывая ходы, и заодно перекусывая, дождевому червю нужно выползать периодически на поверхность, чтобы сбросить с себя землю. При этом, он своим краем хвоста придерживается норы, как бы удерживаясь за неё.
И после, всегда остаются земляные горки. Переработанный червём грунт получается клейким. Замет он высыхает, и делается маленькими, со спичечную головку шариками.
Эти шарики насыщенны витаминами, ферментами, органическими веществами, которые вследствие, убивают в земле все бактерии, предотвращают гниение, что очень важно для корней растений. И еще действуют на состав земли как антисептик, обеззараживая её.
Размножение и продолжительность жизни
Дождевые черви могут быть разнополыми, и гермафродитами. У всех дождевых червей на передней трети части их тела есть утолщения. В них находится яичник и семенник. Гермафродиты друг в друга впускают семя. Уже созревшие яички, в пределах десяти штук, осеменяются. И уползают в разные стороны.
Когда особь женского пола, уже готова к размножению, она сближается с партнёром, совокупляется. На ней образуется что-то вроде кокона, состоящий их нескольких десятков утолщённых сегментов.

Он разделяется своеобразным поясом. В этот кокон поступают все питательные вещества необходимые для расплода. После оплодотворения, червь с себя снимает эту ношу, он попросту соскальзывает с животного.
Края на коконе, с обеих сторон быстро стягиваются, чтобы будущее потомство не высохло, ещё не родившись. Затем, на протяжении четырёх недель созревают и вынашиваются маленькие червячки.
Родившись, они расползаются кто куда. И уже с первых дней своей жизни приступают к активному труду, по переработке земли. А уже в трёх месячном возрасте, подросшие дети достигают размеров взрослых особей.
Ещё один факт о дождевых червях, способность регенерировать. Если кто-то, или что-то разделит его на две половины. Со временем, каждая из половин станет полноценной особью. Это один из способов размножения, но уже не половым путём.

Роль дождевого червя в земледелии очень важна. Во-первых, насыщают почву кислородом, таким нужным для всего растущего на ней. Своими ходами, помогают корням полноценно развиваться.
Равномерно распределяется влага, и почва хорошо вентилируется, взрыхляется. Благодаря постоянному передвижению земли, с помощью червей, из неё извлекаются камни.
Так же, своими переработанными клейкими остатками они склеивают грунт, не давая ему размываться. Ну, и конечно же удобряют землю, когда затягивают в неё листики, личинки насекомых. Это все перегнивает и служит отличными, натуральными био добавками.
1. ОБЩИЕ ЗАМЕЧАНИЯ. НАРУЖНЫЕ ПРИЗНАКИ
Начнем с ознакомления со строением тела дождевых червей. Устройство тела - основа знаний о животных. Хотим ли мы разобраться и разнообразии форм интересующей нас почему-либо группы животных или ознакомиться с образом их жизни, связью их со средой обитания или подойти к решению тех или иных практических вопросов, связанных с этими животными, и т. д. - вопрос о строении тела является основной предпосылкой для решения любых других. В частности, в отношении дождевых червей, уже для того чтобы определить род и вид какого-либо их представителя (а, как мы увидим дальше, их существует немалое число), недостаточно знать его наружные признаки, но необходимо путем вскрытия установить ряд особенностей строения его внутренних органов.
Параллельно мы ознакомимся с работой описываемых органов и их значением в жизни червей.
В теле дождевого червя (рис. 1) можно отличить передний (или головной) конец тела, более толстый, с более сильной мускулатурой и обычно темнее окрашенный, и задний (или хвостовой), более тонкий и более бледный. Задний конец червя часто бывает плоским. На головном конце тела помещается рот, на хвостовом - заднепроходное отверстие. Хорошо отличаются друг от друга также спинная сторона, более выпуклая и обычно более темная, и брюшная - более светлая и более плоская; у червей, консервированных в спирту или в формалине, брюшная сторона может быть вогнутой местами или по всей длине.
Все тело дождевого червя поделено поперечными перетяжками на отдельные участки, которые называются члениками, или сегментами. Эта кольчатость, или сегментация - ведущая черта их организации: каждый из сегментов в принципе имеет одно и то же строение и содержит в основном весь комплекс органов, свойственных этим животным. В передней части тела сегменты более крупные, по направлению кзади их размер постепенно убывает. Число сегментов у обычных видов варьирует в пределах от 90 до 300; оно подвержено значительным колебаниям у разных экземпляров одного и того же вида, но с возрастом в отличие от многих их водных родичей не меняется. Только у некоторых тропических видов число сегментов достигает 600. Внимательно приглядевшись к поверхности тела, можно увидеть, что каждый сегмент подразделен на три части двумя неглубокими бороздками. Это - так называемая вторичная кольчатость, которая также отражает некоторые черты внутренней организации каждого сегмента. Сегменты тела нумеруются, причем первым сегментом считается головной.
Головной сегмент, кроме ротового отверстия, обладает еще одной характерной особенностью: на передней его части имеется головная лопасть - подвижный, меняющий форму придаток, нависающий над ртом. У дождевых червей головной сегмент может быть двоякого рода: либо головная лопасть, вдаваясь на спинной стороне в область первого сегмента, отделена от него поперечной бороздкой, либо она доходит до борозды между 1-м и 2-м сегментами. В первом случае головной сегмент называют эпилобическим, во втором - танилобическим. Эти различия в форме головной лопасти имеют важное значение при определении видов червей (рис. 2).

Головная лопасть - орган осязания и обоняния; ею червь исследует встречающиеся на его пути предметы.
В передней части тела у взрослых особей имеется так называемый поясок, т. е. утолщение, охватывающее от 5 до 12 сегментов, обычно иначе окрашенное по сравнению с остальной частью тела (рис. 3). Кожные покровы в области пояска содержат большое количество желез, выделяющих питательное вещество для яиц при откладке яйцевых коконов. Поэтому в период размножения поясок выглядит сильно набухшим, а тогда, когда откладки коконов нет, область пояска отличается от соседних участков только цветом и иным характером поверхности тела. Форма пояска может быть кольцевой, если он развит одинаково сильно со всех сторон, или седлообразной, если с брюшной стороны он мало развит. По бокам брюшной стороны пояска находятся удлиненные утолщения, которые мы будем называть валиками зрелости (рис. 35). У некоторых видов эти валики заменены несколькими парами бугорков зрелости. Форма, длина, цвет и расположение пояска, валиков и бугорков служат существенными видовыми признаками дождевых червей.
По всей длине тела червя можно заметить маленькие щетинки, которые хорошо видны в лупу. Они находятся на всех сегментах тела, кроме 1-го. У дождевых червей фауны СССР щетинки расположены по 8 на каждом сегменте, попарно или поодиночке. Щетинки образуют с каждой стороны тела червя по 4 продольных ряда, которые принято обозначать буквами латинского алфавита - а, b, с, d (рис. 4). Их расположение имеет большое значение при определении червей. Ряды щетинок а и b, с и d обычно сближены попарно. Степень их сближения у разных видов различна. При определении червей обязательно учитывается также отношение расстояний между рядами щетинок. Эти расстояния обозначаются буквами аа, ab, bc, cd и dd (как принято обозначать отрезки линий в геометрии). Имеет значение также отношение расстояний между щетинками к величине наружного контура поперечного разреза через червя.

Щетинки - важные органы движения: червь может зацепляться ими за частицы грунта или отталкиваться от них при передвижении в почвенных норках и на поверхности земли. Можно также убедиться в их наличии, проведя пальцем вдоль брюшной стороны тела от хвостового конца к головному. Если живого червя поместить на лист бумаги, то будет хорошо слышен характерный шорох при его перемещении, обусловленный трением твердых щетинок. На некоторых сегментах щетинки видоизменены в особые половые щетинки, имеющие значение при спаривании червей.
На брюшной стороне тела, впереди от пояска, помещаются половые отверстия. Сюда относится пара мужских половых пор, обычно находящихся на возвышениях - так называемых железистых подушках (рис. 34) и пара женских половых пор, снаружи плохо различимых.
Кроме того, у большинства видов имеется 2-3 пары пор семеприемников. О значении всех этих отверстий будет сказано далее.
На спинной стороне у консервированных червей хорошо видны в межсегментных бороздах спинные поры, передняя граница расположения которых имеет значение при определении видов червей.
Цвет тела дождевых червей зависит, с одной стороны, от цвета их крови, с другой - от кожных пигментов. Следует строго различать окраску тела червей, о которой можно говорить только в отношении живых особей и которая зависит от комбинации кожного пигмента и цвета крови, от пигментации кожи, которая обусловлена только наличием пигментов. Черви, лишенные пигмента, имеют при жизни розовую или красную окраску тела, а в консервированном состоянии становятся белыми или Сероватыми, пигментированные же виды могут быть красных, бурых, коричневых, желтых и синих тонов.
Длина тела дождевых червей СССР колеблется от 2 до 30 см при толщине от 2 до 12 мы. В тропических странах есть виды, достигающие в длину 3 м. Основная масса червей, населяющих почвы во всем мире, представлена видами, имеющими в длину 5-20 см.
Все дальнейшее изложение относится к дождевым червям семейства люмбрицид (Lumbricidae). Черви других семейств (если не считать ботанических садов, куда черви иногда заносятся вместе с тропическими растениями) могут встретиться только в Уссурийском крае, Средней Азии и в южной части Черноморского побережья Кавказа.
2. ПОКРОВЫ ТЕЛА
Тело дождевых червей покрыто однослойным эпителием. В его составе имеются поддерживающие, железистые и камбиальные клетки (рис. 5).

Поддерживающими клетками осуществляется защитная функция. Наружная часть этих клеток выделяет вещество кутикулы - тонкой прозрачной пленки, покрывающей эпителий. Кутикула состоит из двух систем параллельных волокон, пересекающихся друг с другом под прямым углом. В местах пересечения в кутикуле могут быть отверстия. Направление волокон - диагональное по отношению к продольной оси тела (рис. 6), что наилучшим образом обеспечивает прочность кутикулы при растяжении изнутри (любопытно, что соединительнотканные волокна в коже млекопитающих имеют также диагональное расположение по отношению к продольной оси тела). Кутикула в течение жизни все время снашивается и возобновляется деятельностью эпителия. У консервированных экземпляров кутикула может отставать, и иногда ее можно снять целиком, как чулок.

Кутикулой обусловлена гладкость поверхности кожи, которая облегчает скольжение тела при движении по твердым поверхностям. Она же обусловливает характерную глянцевитость поверхности тела.
Большое значение в жизни червей имеет деятельность железистых клеток. Большая часть их выделяет слизистое вещество, которым всегда смазана поверхность кутикулы; оно выходит на поверхность тела через отверстия в ней (рис. 5 и 6). Это увеличивает легкость скольжения по субстрату и предохраняет тело от высыхания. При любом сильном раздражении слизистые выделения выступают на поверхность тела в огромных количествах: червь мгновенно окутывается толстым слоем густой клейкой слизи. Образование слизистого чехла на теле играет важную роль при спаривании и образовании яйцевых коконов. Кроме того, слизистые выделения покрывают стенки ходов червей внутри почвы, что придает им значительную прочность.
Помимо обычных слизистых клеток, в кожном эпителии дождевых червей имеются на всей поверхности тела так называемые белковые железистые клетки (рис. 5). В области пояска (рис. 25), около щетинок половых отверстий и в других местах тела находятся кожные железы, о значении которых будет сказано далее.
Важным компонентом кожного эпителия являются маленькие клетки, находящиеся в его глубокой части, на границе с подлежащей мускулатурой, и не соприкасающиеся с наружными частя ми поддерживающих и железистых клеток (рис. 5). Это - камбиальные клетки, которые являются резервом; за счет них возобновляются сношенные функционирующие клетки и происходит рост ткани у молодых животных. Эти клетки мобилизуются также при заживлении ран после ранений и иных повреждений.
Из особых клеток кожного эпителия образуются также щетинки. На поверхности тела выступает лишь наружная часть щетинки. Своим внутренним концом она глубоко погружена в стенку тела и может пронизывать ее на сквозь, почти достигая полости тела. Щетинки помещаются в щетинковых мешках, представляющих собой врастания внутрь тела кожного эпителия (рис. 7). Они состоят из вещества, сходного с веществом кутикулы, непрочны и быстро снашиваются. Поэтому в течение всей жизни в глубине щетинковых мешков происходит образование новых щетинок. Каждая щетинка образуется из одной клетки, входящей в состав дна щетинкового мешка.

Щетинки дождевых червей неодинаковы но форме: это палочки, иногда почти совсем прямые, иногда же с явственно загнутыми концами. На некотором расстоянии от наружного конца щетинки находится небольшое утолщение - узелок, т. е. место, к которому прикрепляются мышцы, втягивающие щетинку в глубь тела (мышцы ретракторы; рис. 7). Кроме них, в щетинковых мешках имеются мышцы протракторы, которые одним концом прикрепляются к концу щетинки, а другим - к стенке тела; их сокращением щетинка выталкивается наружу, а также (при неодновременном их сокращении) может совершать довольно разнообразные движения.
О половых щетинках см. ниже (стр. 54).
Говоря о покровах тела, упомянем об интересном явлении свечения дождевых червей, которое с давних пор привлекало внимание многих крупных натуралистов. В частности, о светящихся дождевых червях писал известный исследователь жизни насекомых - Фабр. В разных странах описаны особые виды «фосфорических» червей. Выяснилось однако, что свечение в темноте можно наблюдать и у самых обыкновенных видов. Известный чешский исследователь Вейдовский сообщил, что, раскапывая однажды ночью навозную кучу в поисках дождевых червей, он увидел пятна мерцающего синевато-белого света, который то появлялся, то исчезал в разных точках. Оказалось, что свет исходил от обычных навозных полосатых червей, которых он собрал в большом количестве. Он заметил, что его пальцы после того, как он брал червей, начинали в темноте светиться. Таким образом, светятся слизистые выделения червей, причем только при наличии особых условий, так как свечение наблюдается далеко не всегда. Имеются указания о свечении жидкости, выступающей из ротового и заднепроходного отверстий.
Не может быть сомнения в том, что во всех этих случаях свечение вызвано бактериями, содержащимися в выделениях червей. В процессе жизнедеятельности многих бактерий выделяется световая энергия, освобождающаяся при химических реакциях. Нужно сказать, что почти всегда свечение животных обязано своим происхождением бактериям, так или иначе связанным с ним.
Некоторые исследователи полагают, что свечение полезно для червей: одни думают, что вспышки света содействуют отыскиванию особями друг друга на поверхности земли при спаривании (хотя у червей и нет глаз, они все же способны воспринимать свет поверхностью передней части тела); другие приписывают свечению роль фактора, отпугивающего врагов; третьи думают, что светящаяся слизь, оставляемая червями на их пути, привлекает внимание врагов и делает их самих менее заметными. Однако все это не более чем предположения, не подкрепленные точными наблюдениями.
3. МУСКУЛАТУРА И ДВИЖЕНИЕ. ПОЛОСТЬ ТЕЛА
Основной частью двигательного аппарата дождевых червей является мощно развитая мускулатура их стенки тела (рис. 8). Она устроена но типу так называемого кожно-мускульного мешка. Под кожным эпителием расположен слой кольцевых мышц, сокращение которых уменьшает диаметр червя. Слой кольцевой мускулатуры подстилается слоем продольной мускулатуры (рис. 18), сокращение которой уменьшает длину червя. На границе между этими двумя слоями находится очень тонкий слой диагональных мышечных волокон..

На большей части тела продольный слой мускулатуры имеет значительно большую мощность, чем кольцевой, но в передних 8-12 сегментах тела кольцевой слой может достигать толщины продольного слоя. Эти сегменты играют особенно важную роль при вбуравливании червя в землю (рис. 9).

Прежде думали, что ходы червей в земле образуются поглощением ими земли, т. е. что червь как бы въедается в землю. Однако, как показал уже Дарвин, эти ходы - главным образом результат активной мышечной работы, благодаря которой частицы даже очень твердых грунтов могут быть раздвинуты. Заглатывание грунта во время копания безусловно может происходить, но оно имеет второстепенное значение. Крупным видам дождевых червей, для того чтобы закопаться в плотный грунт на всю длину тела, достаточно 30-40 минут. Эта способность делать ходы в грунте, позволяющая дождевым червям проникать глубоко в землю, подчас на глубину 2 м и более, в значительной мере и определяет космическую роль дождевых червей как почвообразователей. Для этого нужна большая мощность мускулатуры, каковой они и обладают. Мускулатура стенки тела составляет 38-44% от объема тела, а у наиболее сильных видов цифра эта поднимается до 50%. В этом отношении черви из беспозвоночных уступают только пиявкам, у которых мускулатура тела может составлять до 65 % от объема тела.
На поверхности земли и внутри готовых подземных ходов червь, так же как и при закапывании, двигается путем закономерно чередующихся сокращений продольной и кольцевой мускулатуры, комбинируемых с движением щетинок (перистальтические движения). В спокойном состоянии черви передвигаются довольно медленно, но при сильных раздражениях могут очень быстро сокращаться, совершая даже нечто вроде скачков, особенно когда приходится спасаться от преследования. В этих движениях особую роль приобретает продольная мускулатура, содействующая скорости поступательного движения. Черви могут довольно быстро двигаться вверх в вертикальных ходах, сделанных ими в грунте. Опыты в стеклянных трубках с видами родов Lumbricus и Allolobophora показали, что черви опираются спинной поверхностью тела на твердую поверхность трубки. Кроме того, движению червя помогает рот, действующий как присоска (Japp, 1956).
Этим оправдывается не только несравненно большая толщина продольной мускулатуры по сравнению с кольцевым слоем, но и особенности ее строения. У многих видов в продольной мускулатуре наблюдается своеобразная упорядоченность расположения мышечных волокон. Последние укреплены на тяжах соединительной ткани параллельными рядами, благодаря чему на поперечном разрезе они кажутся составленными в «елочку». Такое расположение волокон мускулатуры называется перистым. Оно наблюдается не у всех видов; многим видам свойственно обычное, пучковое расположение волокон продольной мускулатуры.
Для эффективности работы мускулатуры имеет значение то обстоятельство, что под стенкой тела находится полость, наполненная жидкостью. Эта полость сходна по происхождению и характеру с брюшною полостью высших позвоночных и человека, т. е., так же как у них, в ней помещаются внутренности и она выстлана плоским эпителием, носящим название «перитонеальный». У червей полость тела поделена соответственно сегментам тела межсегментными перегородками. Кроме того, полость тела делится на правую и левую стороны брыжейкой, соединяющей брюшную сторону тела с кишечником. В общем, тело червя представляет собой как бы две трубки, вложенные одна в другую: стенка наружной трубки - стенка тела, стенка внутренней - кишечник. Промежуток между ними занят полостью тела, наполненной жидкостью. Все жидкости, как известно, очень упруги и практически несжимаемы при сколь угодно больших давлениях. Поэтому полостная жидкость является антагонистом действия мускулатуры и в известной степени заменяет червю отсутствующий у него скелет. При сокращении мускулатуры стенки тела давление на нее изнутри со стороны полостной жидкости (туpгop) возрастает, и, вследствие несжимаемости ее, поверхность червя приобретает свойства упругого твердого тела. Это помогает ему при движении, а особенно при рытье подземных ходов; передним концом тела червь вбуравливается в грунт наподобие твердого клина.
Упомянем еще раз, что при движениях дождевых червей очень большое значение имеет комбинированное действие мускулатуры стенки тела и щетинок. Особое значение работа щетинок (кроме вбуравливания в землю) приобретает при лазапии по крутизне. Известно, что многие виды червей могут влезать на деревья, что их находят в больших бочках, поставленных для собирания дождевой воды, или в зрелых кочнах капусты под наружными листьями, либо в середине кочна и т. д.
4. КИШЕЧНИК И ПИТАНИЕ
Рот, находящийся на переднем конце тела, ведет в небольшую ротовую полость со складчатыми стенками, за которой следует мускулистая глотка (рис. 10). Благодаря тому, что глотка соединена сложным переплетом мышечных волокон со стенкой тела, она не только совершает глотательные движения и прессует заглатываемые вещества, но может также выворачиваться наружу через широко открытый рот. Эти движения допускают захватывание таких предметов, как листья, камешки и пр., употребляемых для питания или для других целей. В толще глоточной стенки и за ее пределами находятся многочисленные глоточные железы, протоки которых открываются непосредственно в глотку или в особый карман в спинной утолщенной части стенки ее. Глоточные железы выделяют слизистую жидкость, обволакивающую заглатываемые пищевые частицы. В этом отношении их деятельность сходна с деятельностью слюнных желез других животных. Но, кроме того, глоточные железы вырабатывают вещество, переваривающее белки; оно активно в щелочной среде и сходно по своему действию с ферментом, поступающим в кишечник из поджелудочной железы у позвоночных. Таким образом, химическая обработка белков начинается у дождевых червей уже в ротовой полости, что, вероятно, связано с необходимостью возможно более полного извлечения белковых веществ из пищи, как правило, исключительно бедной этими веществами.
Глотка переходит в пищевод (рис. 10). Это довольно узкая цилиндрическая трубка, стенки которой обладают хорошо развитой мускулатурой. По бокам пищевода имеются 1-3 пары боковых карманов (рис. 10) - так называемые известковые железы. У некоторых видов они расположены в толще стенки пищевода и потому снаружи незаметны. Эти железы называются известковыми вследствие того, что в них под микроскопом находят кристаллы углекислой извести. Что эти железы вырабатывают известь - доказывается фактом значительного обогащения ею пищевых масс при прохождении их через кишечник (количество углекислой извести в содержимом кишечника может возрастать от 0.8 до 1.3-1.8%). Предполагалось, что роль этих желез состоит в нейтрализации кислот, содержащихся в заглатываемом грунте. Это предположение стоит в хорошем соответствии с указанной выше необходимостью наличия щелочной среды для деятельности пищеварительных ферментов. Однако вряд ли этим исчерпывается роль известковых желез. Относительно их функции существует много других предположений, притом самых разнообразных; уже это показывает, что функцию известковых желез приходится считать невыясненной.

За пищеводом находится объемистое расширение кишечной трубки - так называемый зоб (рис. 10), занимающий 2-3 сегмента. В нем накопляется заглоченная пища, которая оттуда порциями поступает в следующие отделы кишечника. При отсутствии такого приспособления организм не успевал бы справиться с переработкой поступающего материала. Зоб имеет довольно тонкие эластичные стенки, благодаря чему он хорошо растягивается.
Непосредственно сзади зоба находится другое расширение кишечной трубки - мышечный желудок. Внутри он выстлан эпителием с толстой кутикулой, а стенка его состоит из кольцевого и продольного слоев мускулатуры, причем особенно сильно развит внутренний, кольцевой слой, имеющий «перистую» структуру, наподобие продольного слоя мускулатуры стенки тела. Задача желудка - растирание пищи; в этом процессе главную роль играет, так же как у кур и других зерноядных птиц, трение друг о друга минеральных частиц грунта, между которыми находятся органические пищевые вещества. Дарвин наблюдал, что песчинки и кусочки кирпича, прошедшие через кишечник дождевых червей, принимают вместо угловатой окатанную форму. Имеются новые наблюдения и опыты, доказывающие значение минеральных частиц грунта для перетирания пищи в кишечнике червей; при их отсутствии (например, если поместить червей в торф) они голодают, несмотря на обильный корм в виде листьев (Зражевский, 1953).
За мышечным желудком следует средняя кишка, которая идет до заднего конца тела.
По всей длине средней кишки тянется глубокая спинная складка, или тифлозол, благодаря которой на поперечных разрезах контур полости кишечника принимает подковообразные очертания (рис. 11). Физиологическое значение этой своеобразной черты организации кишечника ясно: таким путем достигается увеличение всасывающей поверхности кишечника. Стенка кишечника содержит большое количество железистых клеток, вырабатывающих слизистый секрет и пищеварительные ферменты. Среди последних, как и в глотке, имеются ферменты, переваривающие белки, и, кроме того, ферменты, превращающие крахмал в сахара (мальтозу и глюкозу); в кишечнике происходит также превращение жиров в растворимое состояние. Таким образом, у червей, как и у позвоночных, питательные вещества в виде растворов всасываются стенкой кишечника. Продвижение пищи совершается действием мускулатуры кишечника, которая состоит из внутреннего кольцевого и наружного продольного слоев мышц (следует обратить внимание на то, что расположение слоев здесь обратное таковому в стенке тела). У некоторых видов имеется несколько слоев мышц в стенке кишечника.

На последних 10-15 сегментах тела кишечник лишен спинной складки, а его эпителий приобретает реснички. Эта часть называется задней кишкой. Всасывания в ней уже не происходит, а только совершается процесс формирования комочков испражнений, т. е. тех копролитов, которые имеют столь важное значение для почвенной структуры. На последнем сегменте тела кишечник открывается наружу заднепроходным отверстием, имеющим вид вертикальной щели.
Интересен спор двух знаменитых натуралистов прошлого века по вопросу о пище дождевых червей - Этьена Клапареда (Франция), превосходного знатока беспозвоночных (в частности, кольчатых червей), и Чарльза Дарвина (Англия). Клапаред нашел в разных частях кишечника дождевых червей остатки измельченных листьев, перемешанных с землей, и на этом основании полагал, что землю черви заглатывают только с той целью, чтобы заглоченные ими растительные остатки лучше растирались. Дарвин, не отрицая того, что черви питаются опавшими листьями и другими растительными остатками, в то же время утверждал, что и заглатываемая земля используется ими для питания. Он наблюдал, что обильно заселены червями и те места, где они могут питаться только землей, богатой органическими веществами (например, аккуратно выметаемые дворы). Все дальнейшие исследования подтвердили правильность наблюдений Дарвина.
Вопрос о способности червей выбирать себе пищу мы затронем в дальнейшем, когда будем говорить о функциях их нервной системы и органов чувств.
Большое значение имеет количество земли, поглощаемой и перерабатываемой в кишечнике дождевых червей. Оно оказалось огромным: взвешиванием копролитов установлено, что черви, населяющие культурные почвы, пропускают через кишечник за 24 часа количество грунта, равное весу их тела.
Для того чтобы закончить рассмотрение кишечника, упомянем о характерной ткани, облекающей снаружи всю среднюю кишку и спинной кровеносный сосуд и заполняющей спинную складку кишечника. При вскрытии живого или только что убитого дождевого червя привлекают внимание желтый цвет и рыхлая бархатистая поверхность средней кишки, на которой контрастно выступают красные кровеносные сосуды. Эта ткань называется хлорагогенной, или желтой. Связь ее с кишечником чисто топографическая: она - измененная часть выстилки полости тела (перитонеального эпителия), прилегающая к кишечнику. Желтая ткань состоит из крупных клеток, плазма которых заполнена каплями веществ, имеющих желтоватую окраску. Происхождение и природа этого вещества, а вместе с тем и функция самой ткани, не вполне ясны. Одни исследователи считают эту ткань местом накопления запасных питательных материалов, наподобие жировой ткани позвоночных. И действительно, включения клеток желтой ткани содержат жир, белок и вещество, сходное с гликогеном (животным крахмалом). В то же время известно, что эта ткань содержит большое количество мочевой кислоты, что инородные вещества, введенные в виде растворов в полость тела (краски), скапливаются в клетках хлорагогенной ткани и что конечные азотистые продукты обмена веществ, подлежащие выведению из организма, обычно имеют желтый или бурый цвет. Все это заставляет думать о выделительной функции этой ткани. Очень вероятно, что наряду с накапливанием запасных питательных веществ клетки желтой ткани имеют способность извлекать из циркулирующей в ней крови и жидкости, заполняющей полость тела, отбросы, образующиеся в процессе обмена веществ. Попадая внутрь клеток желтой ткани, эти вещества выключаются из тока крови и делаются безвредными. Постепенно накапливаясь в клетках этой ткани, они могут длительно там оставаться, но могут и выделяться из организма, так как клетки желтой ткани часто отрываются и попадают в полость тела, а оттуда выводятся наружу вместе с выбрызгиванием полостной жидкости через спинные поры.
5. КРОВЕНОСНАЯ СИСТЕМА. ФУНКЦИИ РАСПРЕДЕЛЕНИЯ ПИТАТЕЛЬНЫХ ВЕЩЕСТВ И КИСЛОРОДА
Распределение питательных веществ, всосанных поверхностью кишечника, осуществляется у дождевых червей при помощи высокоразвитой кровеносной системы. Схема расположения главных сосудов ее следующая (рис. 8, 10 и 12). Вдоль всего тела проходят спинной (над кишечником) и брюшной (под кишечником) сосуды. Спинной сосуд снабжен мускулатурой, которая волнообразными сокращениями гонит кровь от заднего конца тела к переднему. В нескольких передних сегментах (с 7-го по 11-й или, у других видов, с 7-го по 13-й) спинной сосуд сообщается 5-7 парами поперечных сосудов с брюшным сосудом. Эти сосуды снабжены особенно сильной мускулатурой п называются сердцами. Они вполне оправдывают это название, так как служат главным аппаратом, обеспечивающим кровообращение. Кровь, поступающая из сердец в брюшной сосуд, движется по направлению к заднему концу тела. По пути она поступает в сосуды, питающие стенку тела», а также в сосуды, идущие к кишечнику, к выделительным органам (рис. 13), а в соответственных сегментах и к половым органам. Во всех этих частях тела сосуды распадаются на сеть микроскопических капилляров. Из капилляров кровь поступает в поперечные сосуды, которые собирают в конечном счете кровь из всего тела в спинной сосуд.

Имеются и другие продольные и поперечные сосуды, которые можно видеть на рис. 8, 10, 12 и 13; на них мы останавливаться не будем. Особое значение имеет густое сплетение мелких сосудов вокруг кишечника (рис. 13). Сюда поступают питательные вещества, всосанные кишечником, и отсюда они разносятся по всему телу. Заметим, что мускулатурой, хотя и не столь сильно развитой, как в спинном сосуде и сердцах, обладают почти все сосуды, чем предотвращается возможность застоя крови в периферических частях кровеносной системы.

Кровь у дождевых червей, как уже было отмечено, красная. Этот ее цвет обусловлен наличием вещества, очень близкого к гемоглобину крови позвоночных. Однако у червей оно содержится не в тельцах крови, а растворено в жидкой части крови (кровяной плазме). У дождевых червей имеются в крови только бесцветные клетки нескольких типов, в общем тех же, что и тины бесцветных клеток крови у позвоночных.
Как известно, гемоглобин у позвоночных обеспечивает транспорт кислорода из органов дыхания ко всем живым клеткам тела. Ту же роль играет вещество, сходное с гемоглобином, и у дождевых червей. Однако специальных дыхательных органов у них нет: они дышат всей поверхностью тела. Тонкая кутикула и нежность кожных покровов дождевых червей, а также богатая сеть кожных кровеносных сосудов, хорошо обеспечивают возможность поглощения кислорода из окружающей среды. Но отметим, что кутикула дождевых червей хорошо смачивается водой и кислород, по-видимому, у них должен сначала раствориться в воде, смачивающей кожные покровы. Это влечет за собой необходимость сохранения кожи во влажном состоянии. Уже это одно делает ясным, насколько важными для жизни червей являются условия влажности внешней среды. С подсыханием кожи дыхание для них делается невозможным. Однако при попадании и условия недостатка влаги в почве червь может долгое время бороться с этим, используя запасы воды, имеющиеся внутри тела. В этих случаях ему на помощь приходят кожные железы (см. стр. 15), а в случае острого недостатка влаги он утилизирует для этого полостную жидкость, выбрызгивая ее из спинных пор.
Поглощение кислорода поверхностью тела облегчается наличием очень богатой сети кровеносных капилляров, проникающих даже в кожный эпителий (рис. 14) Отсюда кровь через сосуды стенки тела и поперечные сосуды попадает в главные стволы кровяного русла, чем достигается снабжение кислородом всего тела. Красные топа окраски большинства видов дождевых червей (не пигментация, см. стр. 15) определяются именно, наличием богатой сети кожных кровеносных сосудов.

Все это создает возможность жизни червей в условиях очень пониженного содержания кислорода. В этом отношении они приближаются к некоторым из своих отдаленных пресноводных родичей - червям-трубочникам (Tubifex tubifex, Limnodrilus hoffmeisteri и др.), которые, обитая в глубоком илу, могут выносить почти полисе отсутствие кислорода. Относительно дождевых червей есть наблюдения, что они могут жить при содержании кислорода в окружающем их воздушном пространстве, равном 2.5% (как известно, обычно в воздухе его содержится 21%). Даже при наличии 0.4% кислорода в воздухе черви могут поглотить половину количества кислорода, необходимого им для поддержания жизни, и могут находиться в этих условиях довольно долго. Кроме того, попав в бескислородную среду, черви могут переключаться на особый тип обмена веществ, в котором источником энергии для жизненных процессов служат не реакции окисления (для которых и нужен кислород), а распад крахмалоподобного вещества - гликогена, который совершается и в бескислородной среде. Однако запасы гликогена у червей не особенно велики, и, кроме того, при этом способе обмена веществ выделяются кислоты, вредно действующие на организм червей.
Находясь под водой, черви столь же хорошо могут поглощать кислород, как и в воздушной среде. Известно, что они могут жить в воде месяцами при наличии нужного им минимума кислорода и других необходимых для них условий. Этот факт имеет большое значение для понимания многих явлений из жизни дождевых червей.
6. ОРГАНЫ ВЫДЕЛЕНИЯ. ПОГЛОЩЕНИЕ И ВЫДЕЛЕНИЕ ВОДЫ
Функция выделения выполняется у дождевых червей (как у всех кольчецов) трубчатыми органами, расположенными попарно в каждом сегменте, кроме самых передних. Эти органы называются нефридиями, что означает по-гречески «орган подобный почке». Нефридии расположены в полости тела по бокам от кишечника (рис. 8 и 12). Каждый из них представляет собой извитую трубку, начинающуюся внутри тела отверстием в полость тела, расположенным на головчатом расширении, клетки которого снабжены ресничками. Это расширение называют по аналогии со сходными образованиями у более примитивных кольчецов воропкой (рис. 15). Почти непосредственно за воропкой нефридий прободает межсегментную перегородку и проникает в следующий сегмент тела. Там он сначала образует сильно извитую тонкую трубку, которая переходит в более широкую среднюю часть нефридия, снабженную ресничками. Затем нефридий, делая несколько петель, переходит в выводную часть, которая кончается на брюшной стороне тела наружным отверстием (рис. 15), или нефридиальной порой. Снаружи ее найти очень трудно, так как края ее всегда плотно сомкнуты. Недалеко от нефридиальной поры имеется расширение нефридиальной трубки, представляющее собой нечто вроде мочевого пузыря. Нефридии снабжены очень богатой сетью кровеносных сосудов. Кровь, выходящая из нефридия, поступает в поперечный, а из него в спинной сосуд (рис. 16).


Следует заметить, что у одного из дождевых червей (Allolobophora antipae) нефридиальные трубки не открываются независимыми друг от друга порами, а их наружные части впадают в продольные выделительные каналы, которые проходят справа и слева вдоль всего тела и на заднем его конце впадают в кишечник недалеко от анального отверстия. Таким образом, здесь намечается соединение выделительного аппарата в одно анатомическое целое и устанавливается связь с задним отделом кишечника.
Клетки тонкой части нефридиальной трубки улавливают из крови, циркулирующей вне сети нефридиальных капилляров, продукты азотистого обмена, подлежащие выведению из организма. Эти вещества попадают в полость нефридиальной трубки и здесь смешиваются с полостной жидкостью, поступающей через воронку на внутреннем конце нефридия. Полостная жидкость также содержит продукты выделения, отмершие клетки, туда попадают сношенные щетинки и т. д. Жидкость нефридиальной трубки биением ресничек гонится по направлению к выводному концу, откуда периодически выбрасывается через наружную пору сокращением мускулатуры стенки тела (Roots, 1955).
Имеются данные о том, что опорожнение концевого пузыря нефридия происходит один раз в три дня. Другие наблюдения говорят о том, что за сутки червь весом в 1. -1.8 г выделяет 0.82 см3 экскрета. Такие количества его должны выделяться из тела несколько раз в сутки. В экскрете содержатся в общем те же вещества, что и у млекопитающих, а именно: мочевина, аммиак, креатинин, соли и пр., но в значительно более низких концентрациях. Однако нормальный экскрет червей содержит 0.3% белка, тогда как у высших животных белок в продуктах выделения отсутствует.
Клетки средней части нефридиальной трубки обладают способностью к фагоцитозу, т. е. к активному заглатыванию из полости тела нерастворимых в воде веществ (отмершие клетки, свернувшийся белок, бактерии и пр.). Эти вещества накапливаются там неопределенно долгое время. Санитарную службу этого рода несут и другие клетки внутри организма: амебоидные клетки крови, клетки полости тела и вышеупомянутые клетки хлорагогенной, или желтой ткани (см. стр. 26). Особенно много амебоидных клеток, наглотавшихся посторонних телец, встречается в полости тела. Сюда они попадают, активно выползая из сосудов, протискиваясь между клетками сосудистой стенки. Из полости тела эти клетки удаляются разными путями. Во-первых, они проползают через стенку кишечника и, попадая в его полость, выводятся наружу вместе с испражнениями (это наблюдалось много раз); во-вторых, как уже упомянуто, они могут с полостной жидкостью выходить через нефридии и, в-третьих, могут выходить вместе с полостной жидкостью, выбрызгиваемой через спинные поры. Вообще можно думать, что полостная жидкость довольно быстро сменяется. Поэтому она и приобретает у червей такое большое значение в процессе выделения. Роль ее в жизни червей станет яснее после ознакомления с водным режимом их тела.
Мы уже указывали на значение воды в теле червей, когда говорили о роли полостной жидкости (98.8% ее состава - вода) при мышечной работе и о необходимости увлажнения кожи для дыхания (стр. 30). Вода в тело червей непрерывно поступает и выводится обратно в наружную среду указанными выше путями. Таким образом, тело червя и, в частности, полость тела все время прополаскиваются водой. Поэтому для нормального выполнения указанных физиологических функций червям необходимы условия среды, которые обеспечивали бы поступление в их тело воды в значительно большем количестве, чем у большинства наземных животных.
Каким образом вода поступает в тело червей?
Заметим прежде всего, что черви никогда не пьют. Они поглощают воду всей поверхностью тела; вода проходит сквозь покровы и мускулатуру, накапливаясь в полости тела. При этом черви могут использовать воду только в жидком состоянии. Червь в среде, содержащей пары воды, может погибнуть от высыхания, если не имеет иного источника влаги.
При нормальных условиях тело червей содержит около 84% воды. Несмотря на столь значительный запас воды, он оказывается далеко не предельным. Если червю предоставить возможность еще увеличить запас воды в своем теле, он это тотчас же сделает. В этом легко убедиться, если положить дождевых червей в поду. Через несколько часов вес их возрастет на 10-12% за счет воды, всосанной поверхностью тела. После вынимания из воды червь возвращается к исходному весу, причем это происходит за очень короткий срок (1-2 часа). Удаление избыточной воды из организма происходит очень своеобразным путем: она всасывается клетками кишечника, из них поступает в полость последнего и удаляется главным образом через заднепроходное отверстие, а отчасти и через ротовое.
При нормальных условиях жизни в почве функция удаления избытка воды лежит на нефридиях. Наличие постоянного тока поды через организм путем всасывания ее поверхностью тела и выделения ее в избытке почками - явление, весьма обычное среди водных животных. Оно несомненно унаследовано дождевыми червями от их водных предков.
Обмен веществ водных животных совершается при усиленной циркуляции воды через их тело; им не может угрожать недостаток воды, тогда как в почвенных условиях при таком типе водного обмена достаточное количество влаги делается основным фактором, обеспечивающим возможность существования. Поэтому условия влажности почв имеют главное значение в вопросе о заселении их теми или иными видами дождевых червей.
О способности дождевых червей терять воду в периоды засухи и зимовки, что связано с переходом их в состояние скрытой жизни, см. далее (стр. 105).
7. НЕРВНАЯ СИСТЕМА И ОРГАНЫ ЧУВСТВ. РЕФЛЕКСЫ
Вдоль средней линии брюшной стороны тела у дождевых червей под мускулатурой тянется нервный ствол, называемый брюшной нервной цепочкой. В каждом сегменте тела имеется нервный узел, или ганглий, представляющий собой скопление нервных клеток и отдающий от себя 3 пары нервов. Ганглии соединены друг с другом перемычками, коннективами, которые, кроме нервных волокон, также содержат нервные клетки. В переднем конце тела, в 3-м сегменте, брюшная нервная цепочка разделяется на правую и левую глоточные коннективы, образующие окологлоточное нервное кольцо, соединяющееся с надглоточным, или головным ганглием (рис. 17). Этот ганглий - парный, состоит из правой и левой половин, плотно соединенных друг с другом. Но в отличие от всех остальных нервных узлов, лежащих на брюшной стороне тела под кишечником этот ганглий находится на спинной стороне тела и лежит над кишечником. Ганглий этот можно противопоставить всем прочим и вследствие того, что морфологически он сравним с головным мозгом вышестоящих форм (членистоногих). От него идут вперед многочисленные нервные стволы, обильно разветвляющиеся и образующие густые нервные сплетения в первых трех сегментах. Под глоткой, в месте расхождения глоточных коннектив, лежит подглоточный ганглий, представляющий собой результат слияния нескольких ганглиев брюшной нервной цепочки.

Как видно на поперечных разрезах, нервные клетки лежат по периферии ганглия, а средняя его часть занята сплетением отростков нервных клеток (рис. 18). В брюшной нервной цепочке обращают на себя внимание три очень толстых волокна, пробегающих но всей длине тела червя под соединительнотканной капсулой нервной цепочки на ее спинной стороне. Это так называемые нейрохорды, которые еще совсем недавно принимали за гигантские нервные волокна. Однако теперь окончательно выяснено, что они являются своеобразными опорными образованиями (Невмывака, 19476). Эти образования и по строению, и по функции, и по положению между нервной системой и кишечником сходны с хордой позвоночных.

Нервы, отходящие от ганглиев брюшной цепочки, содержат двигательные волокна, заканчивающиеся в мускулатуре, и чувствительные, по которым раздражения поступают в нервную систему с периферии. Тела чувствительных нервных клеток находятся на периферии, в том числе в наружном эпителии (рис. 18). Нервные клетки здесь стоят в ряду эпителиальных клеток. Этот чрезвычайно древний тип соотношений элементов нервной системы сохранился у дождевых червей от их далеких предков, примитивных многоклеточных животных. Очень интересно, что чувствительными нервными клетками здесь становятся не только клетки наружного эпителия, но, как это недавно установлено, также и клетки кишечника, происходящие из внутреннего зародышевого листка (Невмывака, 1947а).
Чувствительные нервные клетки и их окончания имеются и в других местах тела. Ими богато снабжены также нефридии, щетинковые мешки и другие органы. Таким образом, у дождевых червей, как и у высших животных, работа внутренних органов протекает под контролем регулирующей и централизующей роли нервной системы.
Из рефлексов дождевых червей лучше всего известны наблюдаемые в акте ползания. При движении червя по всей длине тела, от переднего конца до заднего, пробегают перистальтические волны комбинированных сокращений мускулатуры. Они следуют друг за другом, и каждая следующая волна может возникнуть еще задолго до того, как первая дойдет до заднего конца тела. Казалось бы очевидным, по аналогии с высшими животными, что причиной этих волн сокращений служит последовательная передача раздражений вдоль по брюшной нервной цепочке. Однако, к удивлению исследователей, выяснилось, что перерезание брюшного нервного ствола и даже вырезание из нее нескольких нервных узлов не останавливают бегущих волн сокращений мускулатуры: волна сокращения проходит через место повреждения так же, как это было и у нормального червя. С тем же результатом можно вдобавок к разрыву брюшной нервной цепочки удалить мускулатуру нескольких сегментов или повредить ее кислотой.
Анализ этих и аналогичных опытов показал, что поступательное движение червя представляет собой длинную цепь рефлекторных актов, в которой каждый сегмент является в значительной мере автономной физиологической единицей. Раздражения с периферии приводят к сокращению мускулатуры данного сегмента. Вследствие этого сокращения раздражаются периферические аппараты в соседнем сегменте, которые вызывают и в нем сокращения и т. д. Таким образом, комбинированные сокращения мышц в каждом сегменте могут представлять собой самостоятельный рефлекс, начинающийся с возбуждения чувствительных клеток на периферии и кончающийся эффектом сокращения мускулатуры этого сегмента. Это наиболее примитивный тип реакции на внешние воздействия. Некоторым его усложнением является передача полученного раздражения по нервной цепочке в соседний задний сегмент тела, в ответ на которое мускулатура этого сегмента сокращается. На рис. 19 изображена схема рефлекса при дугообразном сгибании червя, когда волна сокращений мускулатуры идет по одной стороне тела. Этот рефлекс и является основным при осуществлении поступательного движения червя вперед. Этот способ передачи раздражений по телу, как указано выше, говорит о слабой централизации его нервной системы.

О том же говорят и опыты с удалением надглоточного ганглия. Выше отмечено, что морфологически надглоточный ганглий можно сопоставить с головным мозгом вышестоящих форм (членистоногих). У многих морских кольчецов надглоточный ганглий имеет довольно сложное устройство. Однако у наземных червей надглоточный ганглий подвергся упрощению и физиологическая роль его очень невелика. После удаления надглоточного ганглия удается отметить только некоторое общее расслабление мускулатуры переднего отдела тела, изменения в восприятии света; возможно, он играет роль и при размножении. Но в движениях червя существенных изменений после заживления раны подметить не удается: червь также зарывается в грунт, также избегает опасности и осуществляет все те довольно сложные рефлекторные реакции, с которыми мы ознакомимся далее. Особенно удивительно, что способность к «научению», т. е., по современной терминологии, к условным рефлексам, у червей, лишенных надглоточного ганглия, не пропадает.
Несколько большее значение имеет подглоточный ганглий, так как после его удаления червь лишается многих присущих ему способностей: очень страдают его вкусовые способности (стр. 45).
Было бы ошибочно думать, что слабая централизация нервной системы и относительная автономия отдельных сегментов, обнаруживаемые при поступательном движении червя, означают отсутствие реакций организма как целого. Заранее можно сказать, что таких реакций не может не быть, и, действительно, их очень легко обнаружить. При слабом раздражении заднего конца тела (при легком прикосновении) червь ползет вперед, при раздражении спереди он быстро весь сокращается и ползет в другую сторону, при сильном раздражении в любом месте червь начинает сильно сокращаться дугообразно, в разных направлениях (так называемые гимнастические движения); черви обнаруживают быстрые реакции на свет, на запахи и т. д. Таким образом, указанные выше несовершенство нервной системы и ее слабая централизация обнаруживаются только при внимательном наблюдении и в специально поставленных экспериментах.
Итак, нам известно, что червь располагает довольно богатым арсеналом возможностей для осуществления тех или иных реакций на изменения, происходящие в окружающей их среде.
Рассмотрим теперь, каким путем он может распознавать эти изменения. Средствами для этого являются органы чувств.
Как уже сказано, вся поверхность тела червя покрыта огромным количеством чувствительных нервных клеток. Эти клетки служат органами осязания, развитого у червей очень сильно. Известно, что достаточно, осторожно приблизившись, слабо дунуть на червя, чтобы он ответил резким сокращением продольной мускулатуры; с помощью такого движения он прячется в норку. Вдобавок к чувствительным нервным клеткам в наружном эпителии между клетками имеется очень большое количество свободных нервных окончаний, которые, скорее всего, также обеспечивают функцию осязания.

Как известно уже более ста лет назад, дождевые черви, несмотря на отсутствие глаз, хорошо воспринимают свет. Восприятие света производится особыми светочувствительными клетками, которые по большей части располагаются поодиночке между клетками наружного эпителия (рис. 20). Внутри этих клеток, кроме ядра и густой сети тончайших волоконец - нейрофибрилл, имеется прозрачное светопреломляющее тельце бобовидной или удлиненной формы; его называют линзой по аналогии с хрусталиком глаза более высоко организованных животных. От тела клетки отходит нервный отросток, вступающий в подкожное нервное сплетение и связывающий ее с центральной нервной системой. Такая клетка несомненно представляет собой простейший глаз, как бы изолированную и автономную клетку сетчатки глаза высших животных. Светочувствительные клетки сосредоточены главным образом в передних сегментах тела; больше всего их в головной лопасти, где их может быть свыше 50 (рис. 21). В последующих сегментах число их быстро убывает, в середине тела их не находят, а в последних трех сегментах их снова становится больше. У некоторых видов дождевых червей, кроме изолированных светочувствительных клеток в наружном эпителии, имеются большие группы светочувствительных клеток, расположенные под кожей по ходу нервов, особенно в головной лопасти (рис. 22).


Дарвин тщательно исследовал восприятие света дождевыми червями. Он нашел, что если осторожно освещать их бесшумным фонарем, имеющим только узкий пучок света, интенсивность которого уменьшена красным или синим стеклом (цвет стекла безразличен), то только очень немногие черви реагируют, а именно: уходят в свои норы. Дарвин вел наблюдения над теми видами, которые ночью выходят из нор в поисках пищи или для спаривания; это большой красный червь (Lumbricus terrestris), длинный червь (Allolobophora longa) и некоторые другие. Задний конец у них обычно остается в норке. При более сильном освещении (особенно точные результаты получались при концентрации световых лучей с помощью увеличительного стекла) черви, быстро сокращая продольную мускулатуру, скрываются в норках, «как кролики», замечает Дарвин, приводя выражение одного из своих приятелей, наблюдавшего за его опытами. При этом Дарвин доказал, что реагируют черви именно на свет, а не на лучистую теплоту, исходящую от источника света. Опыты с приближением к червям накаленных кусков железа показали, что они мало чувствительны к лучистой теплоте. Однако, когда черви чем-нибудь «заняты», т. е. когда они втаскивают листья в свои норы, едят и пр., то они не замечают света, даже когда свет концентрировали на них при помощи большого зажигательного стекла. Не реагируют на свет они и во время спаривания. В дальнейшем было доказано, что очень слабый свет может привлекать червей, так как они двигаются по направлению его источника.
Способность ощущать свет играет очень большую роль в жизни червей, так как солнечный свет на них действует губительно (черви очень чувствительны к ультрафиолетовой части солнечного спектра). Реакция ухода в темноту спасает им жизнь (Smith, 1902).
Специальных органов слуха у червей нет. На очень сильные звуки, передаваемые по воздуху, черви не реагируют в том случае, если не вибрирует твердый субстрат, с которым они имеют соприкосновение. Но дрожание твердых тел, с которыми они связаны, вызываемое звуками, они воспринимают очень тонко. Например, по наблюдениям Дарвина, «когда горшок, заключавший в себе пару червей, оказавшихся совершенно бесчувственными к звукам рояля, был поставлен на самый инструмент, то при взятии ноты до в басовом ключе оба моментально спрятались в норы. Через некоторое время они снова показались на поверхности ее, но, когда была взята нота соль в скрипичном ключе, они снова удалились». Эти вибрации крышки рояля воспринимались, очевидно, органами осязания червей.
На высокоразвитом осязании основан метод сбора червей, практикуемый во Флориде: доска или палка втыкается в грунт, обильно населенный червями, и по ее верхнему краю водят другой палкой, как смычком по скрипке (этот способ и называется там «скрипичным»). Пишут, что черви при этом покидают норки и во множестве выходят на поверхность.
В научной литературе издавна от времени до времени появлялись сообщения о звуках, издаваемых дождевыми червями. Действительно, при трении тела и щетинок о грунт, при поршневых движениях во влажных норках, при растирании пищи в глотке, при затаскивании листьев и камешков и т. д. могут возникать звуки. Они тем лучше ощутимы, чем червей больше и чем они крупнее. Но очень сомнительно, чтобы эти звуки имели какое-нибудь биологическое значение.
Кроме чувствительных нервных клеток, нервных окончаний и светочувствительных клеток в наружном эпителии рассеяно большое количество органов, представленных комплексами клеток. Их называют иногда чувствительными почками. Несколько десятков чувствительных клеток образуют цилиндрический или яйцевидный комплекс (рис. 23). Это - чувствительные нервные клетки и длинные нервные отростки, которые идут в брюшную нервную цепочку. Поверхность кутикулы в области чувствительной почки несколько приподнята, а каждая клетка снабжена чувствительным волоском. Эти микроскопические органы распространены в большом количестве по всему телу, но особенно многочисленны они в 1-м сегменте и в его головной лопасти, где у крупных видов их насчитывается около 1800. Функция их точно не установлена. Как полагают некоторые исследователи, часть их может нести осязательную функцию. Но вряд ли можно сомневаться в том, что ими же осуществляются и функции обоняния и вкуса. В пользу такого заключения свидетельствует тот факт, что эти органы имеются в большом количестве в ротовой полости.

Чувство обоняния, т. е. способность к распознаванию различных веществ в газообразном состоянии (что и является способностью воспринимать запахи), развито у червей сравнительно.слабо. В опытах Дарвина черви не реагировали на запах табачного сока, духов, уксусной кислоты, но куски лука (который они очень любят) и листья капусты они находили по запаху. На эфир, поднесенный на близком расстоянии к переднему концу тела, черви реагировали отрицательно и немедленно устранялись от него.
Чувство вкуса, т. е. способность распознавать химические различия веществ при контакте с ними, развито у червей очень тонко и наряду с чувством осязания служит для них главным источником восприятия событий внешнего мира. Опытами Дарвина, развитыми в последнее время рядом исследователей, способность червей выбирать себе пищу доказана совершенно бесспорно и возражения, высказанные некоторыми авторами по этому поводу (например, Тарнани, 1928), несомненно основаны на ошибках.
Очень точная постановка опытов определения вкусовых способностей червей, разработанная Мангольдом (Mangold, 1924, 1951), заключается в следующем. Свернутые в трубку листья вишни или пучок из нескольких сосновых игл перевязывается в нескольких местах нитками и кипятится. Все вкусовые вещества, таким образом, из них удаляются. Затем одна половина такою «испытателя вкуса» окунается в чистый 20%-й желатин, другая половина - в такой же желатин, в который добавлялось испытуемое вещество - измельченные листья разных деревьев и трав, кислоты, хинин и т. д. Такие испытатели вкуса кладутся на ночь на поверхность земли цветочных горшков, в которых культивируются черви. На утро подсчитывают, сколько испытателей черви затащили в норки и при этом отмечают, за какой из концов испытателя ухватился червь. Нужно сказать, что черви, собирая корм, попадающийся им на поверхности земли, никогда его глубоко не вносили в норки, а оставляли его недалеко от наружного отверстия или же только пододвигали к нему. Поэтому вышеуказанный подсчет сделать нетрудно. Если червь не различает концов испытателя вкуса, то при достаточно большом числе повторений опыта должно оказаться, что как за тот, так и за другой его конец червь берется одинаково часто. Если он предпочитает испытуемое вещество чистому желатину, то пропитанный им конец должен оказываться чаще впереди при затаскивании. Наоборот, если вещество оказывается на вкус хуже, чем чистый желатин, то за него червь должен ухватываться реже. Этот опыт видоизменяется подкидыванием червям испытателей вкуса, пропитанных разными веществами, с последующим определением числа втянутых в норки тех и других. Результаты обрабатывались статистически. Проведенные опыты показали, что черви предпочитают гниющие листья только что опавшим осенним; еще менее нравятся им свежие зеленые листья и в еще меньшей степени - высушенные зеленые. Чистый желатин их привлекает более, чем высушенные листья. Гниющие листья разных растений могут быть выстроены в такой ряд в порядке убывающей склонности к ним червей: ива, сладкий лупин, грецкий орех, белая акация, тополь, дуб, горький лупин, липа, бук, клеи, конский каштан. Свежие листья располагаются совсем в другой последовательный ряд. От желатина с примесью хинина черви отказываются, причем чувствуют это вещество уже в концентрации 0.07%. От минеральных кислот они отказываются и любых концентрациях, но прибавка к желатину 1-2% лимонной и фосфорной кислот им нравится. К сахарам они относятся безразлично, но от очень крепких растворов Сахара отказываются вовсе. К сахарину обнаруживается отрицательная реакция уже начиная с ничтожных концентраций.
Способность определять форму тел у червей, по-видимому, отсутствует. Предпочтительное затаскивание ими в норки листьев за передний конец, а сосновой хвои за основание (факт, установленный Дарвином), дальнейшими исследованиями подтвержден. Однако опытами Мангольда установлено, что черви при этом руководствуются только чувством вкуса, позволяющим им отличать верхушку листа от черешка.
Говоря о рефлекторной деятельности дождевых червей, необходимо отметить, что у них давно доказана способность к научению и изменению поведения в связи с пережитыми ранее ощущениями, т. е. условные рефлексы. Не вдаваясь в подробности довольно сложных опытов, установивших этот факт, упомянем, что черви могут «запомнить» дорогу, на которой им не грозит удар электрическим током, а если электрический удар сопровождать прикосновением наждачной бумаги, то черви начинают избегать и наждачную бумагу без электрического тока, хотя она сама по себе и не вызывает изменений в направлении движения червей. В опытах с определением вкусовых способностей червей выяснилось также, что реакция на предлагаемое вещество меняется в связи с предшествующими испытаниями. Черви обычно сначала отказываются от незнакомой им пищи, но потом часто привыкают и берут ее при наличии другой, им знакомой пищи.
Как уже отмечено (стр. 39), аппарат, обеспечивающий наличие условных рефлексов, может быть локализован и в частях нервной системы, не соответствующих головному мозгу вышестоящих форм. Определить, где именно локализована эта функция у дождевых червей, - дело будущих исследований.
Чтобы закончить рассмотрение рефлекторных реакций у дождевых червей, коснемся еще вопроса о болевых ощущениях у них.
Могут ли эти животные испытывать боль?
Замечательный русский зоолог В. Фаусек рассматривал болевые ощущения как полезные приспособления, роль которых состоит в сигнализации наличия повреждений на тело. Он постарался проследить возникновение этого признака в эволюции мира животных и приводит дождевого червя как пример животного, для которого чувство боли еще недоступно. Если мы, укалывая дождевого червя, замечаем его быстрые хлыстообразные движения, то напрашивается аналогия с существом, корчащимся от боли. Насколько необоснованна, однако, такая аналогия показывает следующий простой опыт: если червя, спокойно ползущего вперед, разрезать бритвой пополам, то задняя половина будет хлыстовидно сокращаться, имитируя болевые ощущения высших животных, а передняя будет спокойно продолжать ползти вперед, «не замечая» нанесенного повреждения. Приписать чувство боли задней половине червя и отказать в нем передней - явно нелепо. Но это значит, что и приписывать чувство боли сокращающемуся цельному дождевому червю мы не имеем права.
8. ОРГАНЫ ВНУТРЕННЕЙ СЕКРЕЦИИ
Упомянем о наличии у дождевых червей веществ, вырабатываемых в определенных местах тела и служащих химическими возбудителями разных проявлений жизнедеятельности организма. Такие вещества называются гормонами (греческое слово, означающее «возбуждающий»), а процесс их образования - внутренней секрецией. У позвоночных выработка гормонов происходит частично в специальных железах внутренней секреции (например, надпочечники, щитовидная железа, гипофиз), а также в органах, выполняющих одновременно и другую функцию (например, половые железы, поджелудочная железа, клетки мозга).
У дождевых червей специальных желез внутренней секреции нет, но гормоны вырабатываются в разных частях нервной системы. Уже давно было известно, что в ганглиях брюшной нервной цепочки червей имеются так называемые хромаффинные клетки, которые выделяют адреналин, т. е. вещество, вырабатываемое центральной частью надпочечников высших позвоночных. Это вещество, как известно, является специфическим возбудителем нервных аппаратов, приводящих в движение мускулатуру стенок кровеносных сосудов и служащих важным средством для регулирования ширины просветов сосудов кровеносной системы, а тем самым и кровяного давления. У дождевых червей это вещество играет такую же роль.
В недавнее время обнаружено, что значительная часть нервных клеток надглоточного ганглия также несет внутрисекреторную функцию (Herlant-Meewis, 1956). Различают два типа таких нервносекреторных клеток: одни из них - с однородной протоплазмой, другие - с зернистой. Первые, как полагают, служат регуляторами деятельности половых желез, причем вещество, вырабатываемое ими, по-видимому, тормозит деятельность половых желез: они начинают функционировать в те месяцы, когда заканчивается размножение червей, и исчезают в периоды размножения. Зернистые клетки имеют значение при заживлении ран и восстановлении утраченных частей тела (регенерации): во время этих процессов секреция в них особенно усиливается.
Деятельность пояска дождевых червей, которая заключается в выработке оболочек и питательного содержимого яйцевых коконов, также несомненно регулируется гормонами. Одно время считалось, что гормоны, возбуждающие деятельность железистых клеток пояска, вырабатываются мужскими половыми клетками, созревающими в семенных мешках. Однако это оказалось неверным. Но деятельность пояска несомненно регулируется какими-то гормонами: если пересадить кусок пояска от червя с неактивным пояском к находящемуся в разгаре половой активности, то пересаженный кусок быстро приобретает свойства пояска последнего. Место выработки гормонов, регулирующих деятельность желез пояска, пока неизвестно.
9. ОРГАНЫ РАЗМНОЖЕНИЯ
Дождевые черви размножаются только путем откладки яиц, заключенных в особые яйцевые коконы.
Рассмотрим, как устроены у них органы, обеспечивающие образование яиц, их оплодотворение и откладку. Совокупность этих органов образует половой аппарат Мужские и женские половые органы находятся у дождевых червей в одной и той же особи; таким образом, среди них нет мужских и женских особей, но все они - обоеполые существа, или, как их принято называть, гермафродиты.
Яйца образуются и парс очень маленьких женских половых желез - яичниках, которые укреплены на перегородке между 12-м и 13-м сегментами с брюшной стороны (рис. 24). Яичники устроены очень просто. Они представляют собой комплексы развивающихся яиц; наиболее ранние стадии развития находятся в части, прилегающей к межсегментной перегородке, где яичник состоит из мелких клеток. Наиболее крупные клетки находятся на свободном заднем конце яичника, обращенном в полость тела. Здесь яйцевые клетки достигают своих окончательных размеров (около 0.1 мм в поперечнике) и падают в полость тела. Яйца дождевых червей сферические или слегка вытянутые. Они почти прозрачные, так как в их протоплазме имеется лишь очень небольшое количество зерен питательного материала - желтка. Отсутствие достаточного количества питательного материала для развивающегося зародыша внутри яйца вызывает необходимость снабжения его питанием извне при помощи белка яйцевого кокона.

Яйца заканчивают созревание в так называемых яйцевых мешках. Это - слепые мешковидные выступы межсегментных перегородок, в которые попадают яйца, оторвавшиеся от задней части яичника.
Яйца выводятся наружу через короткие яйцеводы, которые начинаются яйцевыми воронками в 13-м сегменте, затем прободают перегородку между 13-м и 14-м сегментами и открываются на брюшной стороне 14-го сегмента (рис. 24). Яйцевые воронки снабжены ресничками, работой которых яйца улавливаются из яйцевых мешков и в нужный момент (во время образования яйцевого кокона) выводятся по яйцеводу наружу.
Мужские половые железы - семенники - также очень малы. В числе двух пар они помещаются на перегородках между 9-м и 10-м сегментами и между 10-м и 11-м (рис. 24). Мужские половые клетки - сперматозоиды - только начинают развитие в этих крошечных маленьких тельцах. Комплексы будущих сперматозоидов в виде микроскопических комочков округлых клеток падают в полость тела п оттуда попадают в семенные мешки, представляющие собой объемистые разрастания межсегментных перегородок. Количество, форма, расположение и относительные размеры семенных мешков варьируют и служат важным признаком при определении червей.
У некоторых видов дождевых червей (в родах Octolasium и Lumbricus) брюшная часть полости тела возле семенников отгорожена особой стенкой от основной полости сегмента; получаются так называемые семенниковые капсулы. Благодаря их наличию формирующиеся комочки сперматозоидов не могут распространяться по всей полости сегмента и им создается более прямой путь в семенные мешки (рис. 24).
Для выведения сперматозоидов наружу служат семенные воронки и семепроводы (рпс. 24). Семенные воронки обычно крупные; они хорошо заметны при вскрытии червей. Семепроводы, в которые поступают сперматозоиды из семенных воронок, представляют собой очень тонкие цилиндрические трубки, направляющиеся по брюшной стенке тела кзади. Семепроводы от воронок 10-го и 11-го сегментов в 12-м сегменте сливаются друг с другом, и общая трубка семепровода с каждой стороны тела тянется обычно до 15-го сегмента, где переходит через толщу стенки тела и кончается мужским половым отверстием (порой), обычно имеющим вид вертикальной щели.
Мужские половые отверстия сидят на более или менее сильно развитых железистых подушках. Эти подушки, кроме железистых клеток, содержат большое количество сосудов, которые во время спаривания наполняются кровью.
Оригинальной особенностью полового аппарата малощетинковых кольчецов, к которым принадлежат дождевые черви, являются семеприемнники (рис. 24) - небольшие сферические полые мешочки, плотно прижатые к стенке полости тела. Протоки семеприемников проходят сквозь толщу стенки тела и открываются наружными порами, располагающимися в межсегментных бороздах. Стенки семеприемников содержат мускулатуру, действием которой семенная жидкость может всасываться в семеприемник и, наоборот, выбрызгиваться из него наружу. Эта мускулатура действует наподобие резинового колпачка пипетки. Семеприемников бывает 2 или 3 пары; они могут быть расположены сбоку, на брюшной стороне, или могут быть (как в роде Eisenia) сдвинуты на спинную сторону, вплоть до средней линии. Но нужно иметь в виду, что у некоторых видов дождевых червей семяприемники отсутствуют.
К органам, обеспечивающим размножение, следует отнести поясок дождевых червей. У червей, достигших половой зрелости, поясок всегда заметен, но его вид зависит от сезона и состояния упитанности. В периоды размножения поясок сильно набухает. Его функция - формирование яйцевых коконов.
Поясок представляет собой видоизменение наружного эпителия. В области пояска наружный эпителий очень сильно утолщен. Все клетки приобретают железистый характер; среди них можно наметить три типа: 1) относительно мелкие клетки, не содержащие зерен, - слизистые клетки; 2) клетки средней величины, содержащие крупные зернышки, образующие оболочку яйцевого кокона; 3) огромные мелкозернистые клетки, вырабатывающие белковое вещество, составляющее содержимое яйцевого кокона и служащее питанием для развивающихся эмбрионов (рис. 25). Помимо железистых клеток, в пояске можно видеть большое количество кровеносных сосудов и нервных окончаний.

В функции размножения участвует также ряд других желез на брюшной стороне тела между пояском и передним концом тела. Особенно заметны железы на 10-м и 11-м сегментах, которые придают этой части поверхности тела у половозрелых червей беловатый оттенок. Кроме того, около брюшных щетинок в указанной части тела на некоторых сегментах, иногда только с одной стороны тела, бывают развиты железы, заметные в виде небольших припухлостей. Часто сами щетинки изменены, превращены в так называемые половые щетинки, функционирующие при спаривании для удерживания партнера и для раздвигания пор семеприемников. Иногда половые щетинки отличаются от обычных только большими размерами, но у некоторых видов они очень сильно отличаются и по форме (рис. 26). Образуются, с одной стороны, острые стилеты, очевидно вкалываемые при спаривании в кожные покровы партнера, а с другой - щетинки, вводимые в поры семеприемников.

Движение дождевого червя; внешнее строение дождевого червя;
стенка тела, вторичная полость и кровеносная система;
пищеварительная, выделительная и нервная системы; половой
аппарат дождевого червя
Работа 1. Движение дождевого червя. Основную роль в характерном для этих червей движении играет кожная мускулатура. Тело перистальтически сокращается. Укорачивающийся участок становится толще, удлиняющийся - тоньше. Сокращением кольцевой мускулатуры передний участок тела вытягивается, затем начинает сокращаться продольная мускулатура, передний конец укорачивается и к нему подтягивается задняя часть тела.
Вспомогательную роль выполняют щетинки. Свободным своим концом они направлены назад и зацепляются за неровности шероховатой поверхности. Этим путем укрепляется вытянувшийся участок тела и при сокращении мускулатуры подтягивание переднего конца к заднему невозможно из-за препятствий, которые он встречает.
С помощью тех же движений червь зарывается в почву. Передний конец тела вытягивается, утончается и проникает в верхний слой почвы; здесь он, сокращаясь, вздувается, раздвигает почвенные комочки, щетинками фиксируется в щели и, вследствие сокращения, подтягивает сзади расположенный участок тела, В уплотненном грунте дождевые черви строят свои норы путем поедания грунта; пропустив заглоченный грунт через кишечник, они выбрасывают его в виде экскрементов.
Ход работы. Провести беглые наблюдения над движением дождевого червя. Поместить животное на кусок фильтровальной или плотной оберточной бумаги. Ознакомиться с характерным для него типом движения путем перистальтических сокращений тела. Послушать шелестящий звук во время движения, вызванный царапанием бумаги многочисленными щетинками. Провести пальцем вдоль тела червя спереди назад и обратно; многочисленные щетинки, немного наклоненные назад, слегка царапают палец при его движении от заднего конца к переднему.
Работа 2. Внешнее строение дождевого червя. Тело дождевого червя почти цилиндрическое, в поперечном сечении округлое; на заднем конце оно уплощено, на переднем - заметно сужено (рис. 67). Окраска различна, но более или менее темная,
Рис. 67. Дождевой червь: 1 -головной отдел; 2 -анальная лопасть - пигидий; 3 - поясок
буровато-серая, коричневая, красновато-фиолетовая, реже - серая или красная. Спинная сторона окрашена интенсивнее, чем брюшная.
Снаружи хорошо различимы сегменты , или кольца, от которых произошло название типа; в этом суть внешней метамерии тела, в данном случае повторение по длине тела животного однородных колец (рис. 67). Кольца отграничены друг от друга межсегментными бороздками . Не считая деталей строения и размеров, почти все сегменты однородны по внешнему строению; следовательно, сегментация тела гомономная .
На переднем конце тела лежит особая складка округлой формы - головная , или предротовая, лопасть, простомиум . У червей рода Lumbricus срединная (медианная) часть на спинной стороне головной лопасти оттянута назад, так что достигает передней границы второго сегмента; это одна из форм простомиума (танилобичаская головная лопасть; рис. 68). Простомиум дождевых червей лишен органов чувств.
За головной лопастью следует перистомиум , который считается первым сегментом (рис. 68); на переднем крае он несет ротовое отверстие и тем уже существенно отличается от остальных колец. От сегментов туловища отличается по строению и находящаяся на заднем конце тела анальная лопасть-пигидии ; она уступает всем сегментам в размерах и несет на вентральной стороне анальное отверстие (см. рис. 67). Головная и анальная лопасти неравноценны сегментам и таковыми не считаются.
Простомиум, перистомиум и пигидий нарушают гомономный характер сегментации. Вследствие этого тело дождевого червя дифференцировано на три отдела: голову, состоящую из предротовой лопасти и ротового сегмента, туловище и хвостовой отдел (пигидий).

Рис. 68, Передний конец тела дождевого червя. А
- со спинной стороны; Б
- с брюшной стороны (нумерация от 1 до 40 - порядковый счет сегментов); В
- сбоку, правая сторона:
1
- простомиум; 2
- первый сегмент; 3
- ротовое отверстие; 4
- брюшные щетинки; 5
- отверстия семеприемника; 6
- женское половое отверстие; 7
8
- семенная (семепроводящая) бороздка; 9
- поясок; 10
- пубертатные валики; 11
- спинные поры
На спинной стороне тела, на некотором расстоянии от переднего конца (в третьем-четвертом десятке сегментов) у половозрелых особей имеется поясок, клителлум - железистое утолщение покровов нескольких (от 6 до 10) сегментов (рис. 68). У Lumbricus terrestris - на шести-семи сегментах, начиная с XXXI или XXXII по XXXVII включительно (у другого широко распространенного вида-Lumbricus rubellus - поясок ближе к головному концу на 4-5 сегментов). Экскреты железистых клеток пояска играют некоторую роль в процессе размножения. Поясок охватывает тело червя полукольцом, на спинной стороне и боках (рис. 68).
Сегменты вооружены щетинками . Только перистомиум, простомиум и пигидий лишены их. Щетинки расположены на теле метамерно: на каждом сегменте по восемь щетинок, точнее - четыре пары: две пары брюшных и две боковых (или спинных).
Вдоль тела щетинки расположены правильными параллельными рядами. Щетинки слегка изогнуты; большей своей частью они погружены в стенку тела и здесь закруглены; на дистальном (наружном) конце - заострены.
По средней лилии спинной стороны в межсегментных бороздках лежат спинные поры - мелкие непарные отверстия, отсутствующие лишь на нескольких передних сегментах (рис. 68). Через них выступает наружу полостная жидкость, увлажняющая поверхность кожи; возможно, она облегчает скольжение тела по стенке норки и участвует в процессе выделения.
Поясок на брюшной стороне не развит; здесь края его утолщены в виде пары овальных железистых бугорков-пубертатных валиков (рис. 68); с их помощью черви прикрепляются друг к другу при спаривании.
Набрюшной стороне расположены парные отверстия половой системы (рис. 68). Пятнадцатый сегмент несет кнаружи от брюшных щетинок щелевидные мужские половые отверстия, окруженные железистыми утолщениями кожи. Маленькие отверстия женской половой системы лежат (впереди мужских, на четырнадцатом сегменте, также сбоку от брюшных щетинок в непосредственной близости от них. В межсегментных бороздках в области между восьмым и двенадцатым сегментами очень маленькими отверстиями открываются две пары семеприемников. Встречается расположение половых отверстий, отличающееся незначительно от описанного. От мужских половых отверстий до пояска, сбоку от брюшных щетинок, тянется парная семенная , или семепроводящая бороздка , по которой выделяемая наружу семенная жидкость течет к пояску.
Следовательно, передний конец тела до пояска включительно со спинной стороны и вентральной стороны черзя - различны.
Ход работы. 1. Ознакомиться, пользуясь ручной лупой, с внешним видом фиксированного (умерщвленного) дождевого червя; зарисовать передний конец тела со спинной и брюшной стороны; отразить в рисунках сегментацию тела, расположение щетинок, головной отдел, спинные поры, поясок, пубертатные валики, семепроводящие бороздки. 2. Ознакомиться под микроскопом при малом увеличении с внешним видом щетинок на временном микропрепарате; тонким пинцетом извлечь несколько щетинок из тела убитого червя и рассмотреть их в капле воды на предметном стекле.
Работа 3. Стенка тела, вторичная полость и кровеносная система. Стенка тела отгораживает все внутренние органы от внешней среды и служит защитой для них. Пространство внутри от нее занято вторичной полостью, целомом .
Полость тела заполнена жидкостью и разгорожена на многочисленные следующие друг за другом камеры , или сомиты тонкие, прозрачные перегородки между ними - септы , или диссепименты - направлены поперек тела (перпендикулярно главной оси) и сращены со стенкой тела изнутри ее и с поверхностью кишечника (рис. 69). Наличие септ придает полости метамерное строение, выраженное в следовании друг за другом камер целома, Септа образована двумя слоями эпителия с. тонкой мышечной прокладкой между ними. Септы примыкают к межсегментным бороздкам, так что камеры целома соответствуют сегментам наружной кольчатости.

Рис. 69. Передний отдел дождевого червя (вскрыт со спинной стороны), края разреза оттянуты в стороны, показан кишечник; часть его удалена:
1 - 2
- стенка тела (1
- край разреза, 2
- межсегментные бороздки); 3 -
простомиум; 4
- септы (диссепименты); 5
- целом; 6 - 13
- пищеварительная система (6
- глотка, 7
- радиальные мышцы глотки, 8
- пищевод, 9 -
зоб, 10
- мускульный желудок, 11
- средняя кишка, 12 -
посегментные вздутия кишки, 13 -
тифлозолис); 14
- 17
- кровеносная система (14
- сердца, 15
- спинной сосуд, 16
- сосуды стенок кишечника, 17
- брюшной сосуд); 18
- нефридии; 19
- надглоточный ганглий; 20
- брюшная нервная цепочка; I - XXIX - порядковая нумерация сегментов
Кровеносная система . В состав хорошо развитой кровеносной системы отходят продольные, кольцевые и поперечные сосуды, а также сеть кровеносных капилляров.
Два главных продольных сосуда тянутся вдоль тела (рис. 69). Спинной кровеносный сосуд . расположен над кишечником; он обладает собственной мускулатурой и пульсирует. Стенки его ритмически сокращаются и сообщают крови движение. Брюшной сосуд (рис. 70), под кишечником, не пульсирует. Система клапанов ,

Рис. 70. Кровеносная система дождевого червя: А
- схема расположения кровеносных сосудов (вид с левой стороны; капиллярные сосуды не показаны); Б
- схема кровеносных сосудов и кровообращения у дождевых червей на поперечном разрезе (показана сеть капиллярных сосудов; стрелки указывают направление тока крови):
1 - 3
- продольные сосуды (1
- спинной, 2
-брюшной. 3
- субневральный); 4
- 8
- кольцевые сосуды (4
- "сердца", 5 -
спинно-невральный); 6
- спинно-кишечные сосуды; 7
- сосуды стенки тела; 8
9
- сеть капиллярных сосудов в стенке тела; 10
-то же, в стенке кишечника
способных открываться только в одну сторону, пропускает кровь в одном направлении: в спинном сосуде - сзади наперед, в брюшном - в обратном направлении. Вдоль тела тянется еще несколько более мелких продольных сосудов: невральный - под брюшной нервной цепочкой, околонервный парный сосуд, сопровождающий нервную систему справа и слева (рис. 70). Пять кольцевых сосудов лежат в VII-XI сегментах (см. рис. 69 и 70); они метамерно охватывают пищевод и соединяют оба главных продольных сосуда. Кольцевые сосуды пульсируют, за что присвоено им название "сердца́", и гонят кровь, которая внутренними клапанами направляется из спинного сосуда в брюшной. На (всем протяжении тела посегмантно расположены дорзосубневральные кольцевые сосуды , соединяющие невральный сосуд со спинным (рис. 70). К числу метамерных сосудов относятся многочисленные поперечные сосуды ,
отходящие от главных продольных сосудов, но не образующие колец; они связаны с капиллярной сетью (рис.70). Кольцевые и поперечные сосуды на границе со спинным сосудом также имеют клапаны, пропускающие кровь только из кольцевого в спинной. Наиболее тонкие сосуды образуют густое сплетение в стенке кишечника и в поверхностном слое стенки тела; это - сеть кровеносных капилляров , связанная с продольными сосудами через посредство кольцевых и поперечных сосудов (рис. 70).
Система кровеносных сосудов определяет кровообращение (рис. 70). В капиллярной сети, оплетающей кишечник, кровь принимает питательные вещества и продукты обмена; по поперечным сосудам венозная кровь попадает в спинной сосуд. В другой капиллярной сети, разветвленной в стенке тела и органах выделительной системы, кровь окисляется и очищается от конечных продуктов обмена. В связи с отсутствием специальных органов дыхания, газообмен протекает только в коже; увлажнение кожи выделением слизи - необходимое условие для его осуществления. Очищенная кровь по кольцевым (дорзо - субневральным) сосудам попадает в спинной сосуд, где смешивается с венозной кровью.
Таким образом, у дождевых червей кровь движется только по сосудам и с полостью тела не связана, т. е. их кровеносная система замкнутая . Она приняла на себя целиком функцию транспортировки по всему телу питательных веществ от места их образования, освободив от этой функции полостную жидкость. Одновременно обеспечивается газообмен благодаря гемоглобину крови, способному связывать кислород воздуха или растворенный в воде и отдавать его клеткам и тканям животного.
Общее расположение органов кровеносной системы дождевого червя и ход кровообращения с указанием различий их в передней и средней части тела показаны стереоскопически на рис. 71.
Ход работы. 1. Вскрыть дождевого червя. Для этого поместить животное спинной стороной кверху в препаровальную ванночку; вытянуть и прикрепить переднюю часть тела двумя булавками, одной - на уровне первых 3-4 сегментов, в стороне от средней линии, другой - позади пояска, посредине (булавки вкалывать наклонно, отведя головки их в стороны). Лезвием безопасной бритвы сделать неглубокий продольный надрез по средней линии от пояска до переднего конца через всю толщину стенки тела (не задевая кишечник и кровеносный сосуд). Осторожно оттягивая тонким пинцетом края надреза, прикреплять их ко дну ванночки. Струей воды из пипетки промыть препарат. Наполнить ванночку водой. Бегло рассмотреть общий вид вскрытого червя. 2. Подготовить контурный рисунок передней части

Рис. 71. Расположение кровеносных сосудов в теле дождевого червя (блок-диаграмма); справа - область пищевода, слева - область средней кишки: 1
-3
- главные продольные сосуды (1
- спинкой, 2
- брюшной, 3 -
под нервной цепочкой); 4 -
сердца; 5
- боковой; 6 спинно-кишечные; 7
- спинно-невралышй; 8
- сосуды выделительных органов; 9
- сеть капилляров; 10
- сосуды, идущие из брюшного сосуда в стенку тела
тела (1/3 общей длины) для зарисовки по мере изучения внутренних органов; изобразить края взрезанной стенки тела и отметить ограниченную ею полость тела; нанести границы между сегментами. 3. Изучить кровеносную систему на вскрытом животном в той ее части, которая доступна обозрению с помощью ручной лупы; зарисовать спинной кровеносный сосуд, сердца и поперечные сосуды.
Работа 4. Пищеварительная, выделительная и нервная системы. Пищеварительная система состоит из рта, ротовой полости, глотки, пищевода, зоба, мускульного желудка, кишечника и ануса, а также некоторых придаточных органов в виде желез (ом. рис. 69). Ротовое отверстие на брюшной стороне первого сегмента прикрыто сверху и спереди головной лопастью. Ротовая полость , в которую ведет рот, выстлана кутикулой и незаметно переходит в объемистую глотку с утолщенными мускулистыми стенками. В просвет ее открываются протоки многочисленных слюнных , или глоточных , желез , покрывающих глотку извне; секрет их увлажняет и частично переваривает пищу (расщепляет белки). Радиально расположенными мышцами глотка связана со стенкой тела, что придает ей некоторую подвижность (см. рис. 69).
Глотка переходит в узкий трубчатой формы пищевод . начинающийся в VII и заканчивающийся в XIV сегменте. На пищеводе следуют друг за другом кольцевые перетяжки в местах прикрепления к нему диссепиментов. По обе стороны пищевода лежит несколько пар известковых (морреновых) желез , открывающихся в просвет пищевода в передней его части,
Углекислая известь, выделяемая этими железами, нейтрализует содержащиеся в пище кислоты.
Сзади к пищеводу примыкает тонкостенный расширенный зоб, выстланный изнутри кутикулой; роль его с достаточной достоверностью не установлена. За ним следует также расширенный мускульный "желудок" с толстыми стенками, выстланными изнутри тонкой кутикулой (см. рис. 69). В "желудке" при содействии заглоченных песчинок пища перетирается. Ротовая полость, гложа, пищевод, зоб и "желудок" представляют дифференцированные участки переднего отдела пищеварительного аппарата.
Кишечник начинается за "желудком" и заканчивается почти у ануса. В местах прикрепления диссепиментов кишечная трубка образует кольцевые перетяжки. Между перетяжками метамерно следуют посегментно-расположенные расширения. В просвет кишечника выделяются клетками его стенок разнообразные ферменты; в других клетках кишечника происходит всасывание переваренной пищи. Короткая задняя кишка заканчивается анальным отверстием.
Выделительную систему образуют многочисленные парные органы - метанефридии , посегментно расположенные в полости тела (см. рис. 69). Небольшая воронка метанефридия - нефростом обращена в полость тела; края ее усажены мерцательными ресничками (рис. 72). От нефростома отходит узкий канал, пронизывающий септу; в полости следующего кзади сегмента он образует несколько петель и открывается наружу выделительным отверстием - нефрoпорой . Нефридиальный канал снабжен ресничками и погружен в особую оболочку - тело метанефридия, состоящее из целомического эпителия, пронизанного густой сетью кровеносных сосудов (см. рис. 72). Отбросы (мочевина, аммиак), образующиеся в клетках тела в результате расщепления сложных органических соединений, извлекаются стенками нефридиальнсго канала из крови в капиллярной сети почки; согласованным действием ресничек целомическая жидкость, а вместе с ней и экскреты, гонятся к выводному отверстию (нефропоре).
Нервная система состоит из головного мозга и брюшной нервной цепочки (рис. 73). Головной мозг, т. е. парный хорошо развитый надглоточный ганглий , помещается на спинной стороне глотки в переднем ее конце. Отходящая от него парная мощная коннектива связывает его с

Рис. 72. Выделительная система дождевого червя. А
- метанефридий; верти, кальные линии изображают диссепимеиты; по одну сторону септы расположена воронка - нефростом, по другую извитой выделительный канал (зачернен), стрелки в канале и в нефростоме указывают ток жидкости с экскретами, стрелки в иных направлениях - ток крови в толще почки; в ней. показана часть кровеносной системы; Б
- нефростом, вид с брюшной стороны; В
- тоже, продольный разрез:
1
- 6
- воронка-нефростом (1
- внешний вид, 2
- клетки мерцательного эпителия, 3 -
реснички, 4
-- входная щель, 5-"нижняя губа", 6
- "верхняя губа"); 7
-- 10
-выделительный канал (7
- начало канала, идущего от воронки, 8
- петли выделительного канала, 9
- мочевой пузырек, 10 -
нефропора); 11
- ткань целотелия
подглоточным ганглием, охватывая глотку с обоих боков. В целом зги элементы центральной нервной системы образуют окологлоточное кольцо (рис. 73). Брюшной нервный ствол, началом которого служит подглоточный ганглий, тянется вдоль тела на брюшной стороне (рис. 73). В каждом сегменте нервные клетки сконцентрированы, образуя парное скопление, или ганглий ; ганглии смежных сегментов связаны парной коннективой. В целом образуется брюшная нервная цепочка . Хотя ганглии не резко обособлены, мета мерность строения нервной системы выступает вполне отчетливо. Каждый ганглий иннервируег свой сегмент: от него отходят нервы к различным органам в том же сегменте.
Таким образом, метамерия присуща внешнему строению, полости тела, пищеварительной, выделительной и нервной системам, так что тело дождевых червей представляется состоящим из следующих друг за другом однородного строения участков, или метамер. Наружная сегментация и метамерия внутренних органов совпадают.

Рис. 73. Нервная система переднего конца тела дождевого червя. А
- вид со спинной стороны; Б
- вид сбоку (левая сторона):
1
- парный надглоточный ганглий - "мозг"; 2
- 3
- нервы, отходящие к простомиуму и I сегменту (2 - ветвь простомиума, 3
- ветвь I сегмента); 4
- окологлоточная коннектива; 5 - брюшная нервная цепочка; 6
- ганглии, посегментно расположенные; 7
- рот; 8
- глотка; 9 -
пищевод; 10
- предротовая лопасть; I-VI - сегменты тела
Описанные порознь внутренние органы дождевого червя размещены определенным образом; внешний их вид и внутренние связи между ними (топография) изображены стереоскопически на рис. 74,
Ход работы. 1. Изучить с помощью ручной лупы и зарисовать передний отдел пищеварительной трубки и часть средней кишки; отметить функции органов пищеварительной системы; убедиться в сращении кишечника с септами. 2. Ознакомиться с выделительной системой, ее органами и их метамерным расположением, пользуясь ручной лупой; приготовить временный микропрепарат; пинцетом вырвать один метанефридий и рассмотреть его в капле воды под микроокопом при большом увеличении. 3. Изучить нервную систему; отыскать на глотке парный

Рис. 74. Стереограмма отрезка тела дождевого червя; два сегмента вскрыты со спинной стороны, края разреза развернуты в стороны; представлена топография внутренних органов:
1
- стенка тела; 2
- септы; 3
- камеры целома; 4 -
кишечник; 5
- его тифлозолис; 6
- сегментарные органы - метанефридии; 7 - 11
- кровеносная система (7
- спинной продольный сосуд, 8
- брюшной сосуд, 9
- субневральный, 10 -
боковой невральный, 11
- поперечный спинно-кишечный сосуд); 12 -
брюшная нервная цепочка
беловатый надглоточный ганглий; удалить кишечник, перерезав глотку, и найти брюшную нервную цепочку.
Работа 5. Половой аппарат дождевого червя. Половая система дождевого червя (рис. 75) -гермафродитная. Она сосредоточена в немногих сегментах переднего конца тела. Мужские гонады - семенники лежат в X и XI сегментах, яичник - в XIII сегменте.
Три пары мешковидных семенных пузырей расположены: передние, наименьшего размера, IB IX сегменте; вторая пара - в XI и последняя, самая крупная - в XII сегменте; все они образованы разрастанием диосепиментов соответствующих сегментов. В две непарные цилиндрической формы семенные капсулы открываются семенные пузыри; первая и вторая пары открываются в переднюю капсулу (в X сегменте), а третья пара - в заднюю (в XI сегменте). Стенки капсулы образованы разрастанием эпителия, выстилающего целом, а полость капсулы есть часть самого целома (рис. 75).
В семенных капсулах по бокам брюшной нервной цепочки лежат очень маленькие гроздевидные семенники (длиной менее одного миллиметра), прикрепленные попарно к передней стенке камер целома X и XI сегментов. Против каждого семенника на задней стенке той же целомической камеры сидит по паре более крупных органов - половых воронок , сильно

Рис. 75. Половая система дождевого червя. А -
схема строения половой системы у животного, вскрытого со спинной стороны. На правой стороне семенные мешки удалены; правая половина семенных капсул вскрыта для обнаружения семенников и половых воронок; Б
- схематичный продольный разрез через сегменты, содержащие половой аппарат:
1-3
-стенка тела (1
- наружная поверхность, 2
- внутренняя поверхность, граница полости тела, 3
- сегменты); 4
-диссепименты; 5
- передняя и .6
- задняя семенные капсулы; 7
- семенники; 8 -
мужские половые воронки; 9
- семепротоки; 10 -
семепровод; 11
- мужское половое отверстие; 12
-14
- семенные мешки, или пузыри (12
- передний, 13
- средний, 14
- задний); 15
- яичник; 16
- женская половая воронка; 17 -
женское половое отверстие; 18
- семеприемники
складчатых и снабженных ресничками (рис. 75). Семенники не поддаются обнаружению, если половые клетки из них уже переместились в семенные пузыри.

Семепротоки (семевыносящие каналы) от каждой воронки направляются назад, сливаются на каждой стороне тела в общий семепровод; оба семепровода, пробегающие под целомическим эпителием, также не видны. Они тянутся до XV сегмента, где открываются наружным парным мужским половым отверстием.
В состав женской половой системы входят яичники, яйцеводы и семеприемники (рис. 75). Парный яичник расположен в XIII сегменте; это небольшие грушевидные органы, прикрепленные к переднему диссепименту по бокам брюшной нервной цепочки. На заднем диссепименте того же сегмента (против соответствующего яичника) сидят две женские половые воронки . Короткий парный яйцевод проходит сквозь диссепимент и в следующем (XIV) сегменте открывается парным женским половым отверстием. Две пары семеприемников в форме небольших шаровидных мешочков лежат попарно в IX и X сегментах.
У половозрелых особей сперматозоиды поступают из семенников в семенные мешки (через капсулы), где они дозревают. Через половые воронки они движутся по семепритокам и семепроводам к наружному половому отверстию. При копуляции черви прикладываются друг к другу брюшными сторонами передних концов тела так, что поясок одного приходится против отверстий семеприемников (IX и X сегменты) другого (рис. 76). Семенная жидкость, выступающая из мужского полового отверстия (на XV сегменте), гонится по семенным бороздкам на брюшной стороне тела вплоть до пояска и здесь засасывается семвприемниками другой особи. Сперма сохраняется в семеприемниках и используется позднее для оплодотворения яиц. После копуляции яйцеклетки по выходе из яичника попадают в женскую половую, воронку и по короткому яйцеводу - наружу. Здесь они встречают особую слизистую массу, охватывающую тело кольцом и формирующую кокон; она вырабатывается эпителием
9 - субневральный, 10 - поперечные спинно-кишечные); 11 - 13 - брюшная нервная цепочка (11 - окологлоточное кольцо, 12 -_брюшная цепочка, 13 - ее ганглии); 14 - простомиум; 15 - кожные покровы; 16 - 17 -мускулатура; 18 - 22 - передняя кишка (18 - ротовое отверстие, 19 - глотка, 20 - пищевод, 21 - зоб, 22 - мышечный желудок); 23 - половая система
пояска. Слизистое кольцо, сползая с тела через передний конец, скользит мимо отверстий семеприемников, из которых выступают сперматозоиды, оплодотворяющие яйцеклетки. Кокон развивается в почве.
Общий вид расположения внутренних органов, включая половую систему, дан на рис. 77.
Ход работы. Рассмотреть половую систему дождевого червя. Раздвинуть в стороны семенные мешки (или семенные пузыри), сосчитать их Число, рассмотреть форму и расположение в сегментах; найти семенные капсулы. Удалить семенные мешки на одной стороне тела, глазными ножницами осторожно вскрыть семенные капсулы, промыть полость капсул пипеткой, найти семенники и семепрнемники (семепроводы, пробегающие под целомическим эпителием, не обнаруживаются). Зарисовать контуры передней части тела (18 сегментов) вскрытого червя и (пользуясь отчасти рис. 75) органы полового аппарата.




